El fenómeno alelopático. El concepto, las estrategias de estudio y su aplicación en la búsqueda de herbicidas naturales.
Alberto de J. Oliveros-Bastidas
Universidad de Los Andes – Facultad de Ciencias
Departamento de Química – Grupo de Química Ecológica
Núcleo Universitario Pedro Rincón Gutiérrez – Mérida 5101-A
Venezuela
e-mail: aloliver@ula.ve
RESUMEN.
Las plantas poseen como característica general en ser sedentarias, desarrollándose en el mismo lugar donde ocurre la germinación. Este hecho limita a las plantas del escape, el principal mecanismo de defensa en la naturaleza. Sin embargo, esta limitación ha sido en parte compensada con otros mecanismos de defensa, como lo es la comunicación con su entorno, el cual le permite detectar agentes que le pueden ser perjudiciales. Dentro de los sistemas de comunicación, aquella mediadas por agentes químicos es la que más ha sido estudiada, y en especial aquella que tiene lugar ente diferentes especies, donde la participación de un agente químico es el responsables de controlar tal comunicación. Este fenómeno es conocido como la alelopatía, y los agentes causantes de la comunicación son denominados aleloquímicos. En este trabajo se lleva a cabo una revisión de los conceptos introducido para definir el fenómeno alelopático, las diferentes estrategias o metodologías para estudiar en condiciones de laboratorio la operatividad de este fenómeno como una evidencia de la existencia del fenómeno, así como la aplicación de las estrategias de estudio en la búsqueda de moléculas novedosas para el desarrollo de herbicidas.
Palabras clave: Alelopatía, aleloquímico, bioensayos, herbicidas.
ABSTRACT.
A general characteristic of the plants is to be sedentary, i.e. to grow up at the same place of their germination. This fact limits the plants to escape from attacks, which is the main mechanism of defense in nature. However, this limitation has been partly overcome using other mechanisms of defense, such as chemical communication with its environment. This phenomenon is known as allelopathy, and the agents in charge of that communication are called allelochemicals. The aim of this paper is to review the concepts introduced to define the allelopathic phenomenon, the strategies and methodologies to study it in the laboratory, as well as, research strategies in the way of searching new molecules for the development of natural herbicides.
Key Words: Allelopathy, allelochemicals, bioassay, herbicides.
Introducción
Una característica notable de la mayoría de las plantas terrestres es que en su estado adulto son sedentarias. Aunque algunas especies de plantas pueden desarrollarse vegetativamente en diferentes direcciones desde su lugar de anclaje, la mayoría permanecen en el mismo lugar de su germinación. Esta existencia sedentaria tiene un profundo impacto en la historia de vida de los individuos, causando una influencia en la ecología y evolución de la población entera. La germinación, el establecimiento de la plántula, su crecimiento, su reproducción y forma de dispersarse; son afectados de manera notable por las características del medio de crecimiento. Por ejemplo, la cantidad y composición de la luz incidente, modificada drásticamente por el micro ambiente de la planta, o las sustancias liberadas por las plantas vecinas, pueden afectar la germinación de las semillas y el establecimiento de la plántula.
Así, las plantas que están sujetas a desarrollarse en el mismo lugar de su germinación experimentan competición provenientes de sus vecinos de una manera más agresiva que en el caso de los animales, donde este sedentarismo y anclaje son menos acentuados, y otros tipos de defensa como el escape, la evasión y el mimetismo, son las que se han perfeccionado evolutivamente.
Esta característica de un restringido de movimiento en las plantas, ha sido compensada evolutivamente con otros mecanismos de defensas, subdivididas en mecánicas, fenológicas y químicas (1-3). Entre las mecánicas encontramos la formación de espinas, aumento en la dureza de sus hojas mediante una mayor deposición de sílice y presencia de tricomas entre otros; las cuales afectaran la morfología de la planta. Entre las defensa fenológicas encontramos por ejemplo, una rápida reposición de biomasa perdida y “escape fenológico”, definido como el ajuste del ciclo biológico de las plantas para evitar condiciones de estrés o más favorables, para disminuir la disponibilidad de las plántulas cuando los herbívoros son más activos. Por último, las defensa de tipo química (constitutiva o inducida), están representadas por aquellos compuestos de origen secundario que actuaran en detrimento del estrés ocasionado a la planta, como por ejemplo, contra la herbívoría, áfidos o patógenos.
Todas ellas, de manera individual, aditivas o sinérgicas, le han aportado a las planta una excelente capacidad adaptativa, confiriéndolas de una alta prevalencia para algunas especies, en diferentes nichos ecológicos. Las defensas de tipo químicas son metabolitos secundarios, responsables de la transmisión de información de las plantas con su entorno, e incluyen la interacción planta-planta, planta-insectos y planta-microorganismo.
Dependiendo del tipo de interacción o información a transmitir, muchos de estos metabolitos son sintetizados desde los primeros estadios de crecimiento (defensa constitutiva), como en el caso de las interacciones planta-planta de la misma especie, o por el contrario, estos pueden ser sintetizados por estímulos externos, como la presencia de herbívoros, que se traducen en una alarma metabólica que disparan o catalizan sus rutas biosintéticas (defensa inducida) (4).
Siendo estos metabolitos secundarios responsables de la interacción entre las plantas con su entorno, no es sorprendente que hayan sido asociados a estos compuestos una amplia variedad de actividades biológicas, medidas en ensayos in vitro o in vivo (5). El estudio de la dinámica de estas defensas químicas, comprende dos aspectos fundamentales. Primero, la observación del fenómeno en su hábitat natural y en segundo lugar, debido a la gran variedad y complejidad de variables (6-8) llevar a nivel de laboratorio el estudio del efecto previamente observado.
Es este control permitido a nivel de laboratorio el que ha logrado evaluar, de una manera separada, la interacción de los diferentes fenómenos operantes en las interacciones de las plantas con su entorno (9-16). Dos de estos fenómenos son la competencia y el fenómeno alelopático entre plantas (10). Sin embargo, aunque es posible separar y estudiar la interacción alelopática de manera independiente, la determinación de la importancia relativa de estos fenómenos ha sido muy difícil, debido principalmente a las dificultades experimentales encontradas al separa sus efectos (17,18).
Desde el punto de vista ecológico, existe una necesidad imperante de definir con mayor precisión los mecanismo en ambos fenómenos (19-21), siendo escasos los trabajos que han logrado dar peso específico a ambos fenómenos (22), dentro de los cuales se ha demostrado que compuestos constitutivos de exudados de la raíz son los responsables de efecto alelopático (23), y el balance entre ambos fenómenos fue regulado, según la disponibilidad de exudados en el medio.
Básicamente, el fenómeno alelopático implica un componente ecológico (la evidencia de que exista en la naturaleza), un componente químico (aislar, identificar y caracterizar los aleloquímicos) y un componente fisiológico (interferencia en los procesos bioquímicos o fisiología, tanto a nivel celular como molecular). Y todo ellos han de ser abordados para estudiar la implicación del fenómeno alelopático en un determinado ambiente.
El concepto del fenómeno alelopático. Una revisión.
La definición más tradicional del fenómeno de alelopatía, es la postulada por Rice (24), descrito como “cualquier efecto directo o indirecto causado por una planta (incluyendo microorganismos) sobre otras a través de la producción de compuestos químicos que escapan al medio ambiente”. La definición más amplia, es la desarrollada por la Sociedad Internacional de Alelopatía en 1996 (25), definiéndola como: “cualquier proceso que involucre metabolitos secundarios producidos por plantas, algas, bacterias y hongos, que influyan en el crecimiento y desarrollo de sistemas biológicos y agrícolas”.
Dependiendo de cada sistema y de las condiciones de trabajo, han sido introducidas otras definiciones para describir el fenómeno, como por ejemplo, en los estudios realizados por Inderjit y Dakshini (26) en sistemas acuáticos, definiendo la alelopatía como “un fenómeno donde las alomonas generadas por las algas pueden afectar 1) otra alga en su vecindad, 2) su propio crecimiento, 3) la asociación con los microbios, 4) plantas superiores de su vecindad y 5) acumulación y disponibilidad de iones nutrientes; los cuales influyen sobre la distribución, crecimiento y establecimiento de otras algas, microorganismos y plantas”.
A diferencia de las dos anteriores, esta representa una definición más precisa y detallada del sistema al que hacen referencias (sistemas acuáticos). Así como esta definición, han sido descritas otras, i) “ mecanismo de interferencia mediante el cual una planta libera una sustancia que afecta a otra planta” (27); ii) “el efecto negativo de una sustancia liberado por una especie de planta sobre el crecimiento y reproducción de otra” (28); iii) “la liberación de compuestos extracelulares que inhiben el crecimiento de otros microorganismos” (29); iv) “supresión del crecimiento de la planta vecina por la liberación de compuestos tóxicos” (30); v) “la liberación de compuestos químicos por un invasor que tiene efectos perjudiciales sobre los miembros de la comunidad de plantas hospederas” (31); y vi) “la supresión química de especies de plantas competidoras” (32).
A partir de estas definiciones es posible extraer tres rasgos imprescindibles a ser tomados en cuenta al estudiar la existencia del fenómeno alelopático: I) la liberación de un compuesto al ambiente encargado de transmitir un efecto, II) la absorción por el organismo receptor y III) provocar un efecto sobre su normal crecimiento. La complejidad del fenómeno resulta de las variables nacientes de cada uno de estos procesos, como por ejemplo, el mecanismo de liberación del aleloquímico y su estabilidad en el ambiente, que definirán los niveles y la forma en que lo absorberá el organismo receptor, y los innumerables modos de acción encontrados, muchas veces dependiente del ente receptor.
Sorprendentemente, como hemos referido, muchos de estos procesos han sido controlados a nivel de laboratorio y a nivel de campo, lo que ha dado alta factibilidad de operación del fenómeno alelopático, como es el caso de (±)-catequin y la introducción de plantas invasoras no nativas, lo que será reseñado más adelante. Sin embargo, la forma más simple de evidenciar el fenómeno alelopático, es la interacción de dos plantas y el registro del agente alelopático desde la planta donadora a la receptora, y la observación de algún efecto sobre las condiciones normales de su desarrollo.
A pesar de esta simplicidad, no existen datos que demuestren la traslocación de un aleloquímico de su planta productora a la planta receptora, utilizando plantas vivas e intactas en los dos casos. Para las benzoxazolinonas, uno de los aleloquímicos con más amplias actividades biológicas, varios diseños experimentales utilizando cocultivos de las plantas donadoras y receptoras, han demostrado que existe una correlación directa en los niveles de benzoxazolinonas medidos en los exudados de la raíz, y el efecto observado sobre el desarrollo de diferentes plantas receptoras.
Sin embargo, a la fecha no se encuentra en la literatura diseños que hayan logrado el seguimiento del aleloquímico en todo el sistema, esto es, la planta donadora, el medio de crecimiento, planta receptora y algún efecto en sus parámetros de crecimiento, que en resumen, son los principios básicos del fenómeno anteriormente señalados.
La ruta de liberación de los aleloquímicos
El estudio del fenómeno alelopático es complejo, pues existen diversos factores involucrados, especialmente el trabajo con sistemas vivos, así como la necesidad del empleo de diversas técnicas instrumentales, convirtiendo al estudio de la alelopatía en una ciencia multidisciplinaria. Sin embargo, cualquiera que sean las propiedades de los compuestos detectados en la planta, las características tales como actividades biológicas diversas, tiempo de vida media en el suelo, especificidad a un restringido grupo de organismos, ser inocuo para la especie donadora; evitando de esta manera fenómenos de auto toxicidad; son las exigidas para se considerado como un aleloquímico.
No obstante, ninguna de estas característica tendrán algún significado ecológico, si este aleloquímico no es liberado al ambiente por un mecanismo lo suficientemente efectivo para garantizar un nivel de concentración toxico a la planta u organismo receptora. Ecológicamente, se han propuestos cuatro mecanismos para la excreción de los aleloquímicos al ambiente, involucrando fenómenos de volatilización, lixiviado, descomposición de partes de la planta en el suelo y exudados por raíz. Estos mecanismos implican que los aleloquímico pueden encontrase en cualquier parte de la planta, y que la dinámica de su actividad es una función del mecanismo en que este pueda ser liberado. Por ejemplo, la descomposición de residuos vegetal implica material senescente, que no implica una función activa de la planta, mientras que volatilización y exudados de raíz, son procesos extremadamente activos donde está involucrado el tejido vivo de la planta.
Estrategias aplicadas en el estudio del fenómeno alelopático
El estudio del fenómeno alelopático en la naturaleza, como se ha indicado, implica en primera instancia, la observación de los rasgos microscópicos del sistema, y a partir de esta observación, tratar de discernir parámetros poblacionales que indiquen un dominio de una especie sobre otra, tales como predominio, la exclusión y la distribución de las especies vegetales existentes, como indicadores del dominio poblacional de una de las especies que constituyen el ecosistema (9-16).
Sin embargo, estos son sólo parámetros macroscópicos, es la manifestación de una serie de procesos que a nivel microscópico ocurren en el sistema, que denominamos el componente químico, y que es el responsable de las alteraciones bioquímicas y fisiológicas que causan los rasgos macroscópicos observados. Es a partir de este componente químico (aleloquímico), y la separación de las diferentes interacciones que pueden estar operando, tales como la alelopatía y la competencia por nutrientes (33), lo que representan las principales limitaciones de los resultados, y los orígenes del escepticismo manifestado por muchos ecólogos (34).
Los primeros estudios que abordaron el efecto de las condiciones ambientales sobre el desarrollo de las plantas se llevaron a cabo estudiando variables abióticas, como condiciones nutricionales, presión parcial de CO2, alcalinidad y alteraciones de las propiedades fisicoquímicas del suelo (35, 36).
Sin embargo, las evidencias iniciales del efecto de una planta sobre otra, data del año 300 antes de cristo, basada en la observación del comportamiento macroscópico de las comunidades de plantas, donde se observó que muchas cosechas, tales como leguminosas y gramíneas, destruían muchas malas hierbas, e inhibían el desarrollo de otras cosechas (37).
Estas primeras observaciones sobre interacción planta-planta fueron realizadas por el botánico griego Theophrastus en las que demuestra el valor de la observación de los rasgos macroscópicos, enunciando la siguiente conclusión:
“Nosotros debemos considerar el carácter distintivo y la naturaleza general de las plantas, desde el punto de vista de su morfología, su comportamiento bajo condiciones extremas, su modo de generación y el curso entero de su vida” (38).
En cierto sentido, estas conclusiones siguen siendo válidas, y son aplicadas en los diseños experimentales para el estudio de la alelopatía, donde el termino “comportamiento bajo condiciones extremas” y “curso entero de su ciclo de vida”, constituye el término “tratamiento” y “respuesta”, en la mayoría de los dispositivos experimentales diseñados para aislar el efecto alelopático de otros fenómenos en las interacciones de la planta con su entorno.
Sustentado en los términos tratamiento y dosis, los experimentos en el campo de la alelopatía inicialmente estaban basados según los postulados del bacteriólogo Alemán Robert Coch -1843-1910- (39), donde cita una serie de condicionantes que necesitan ser totalmente establecidas para asegurar que un organismo es causante de una enfermedad, resumidas en cuatro directrices:
1.- La enfermedad tiene que estar siempre asociada a la presencia de un patógeno.
2.- El patógeno debe ser aislado del tejido enfermo in vitro.
3.- El patógeno inoculado sobre un organismo sensible, debe manifestar los mismos síntomas originalmente encontrados.
4.- El patógeno es re aislado del tejido enfermo, y debe ser idéntico al inoculado.
Así, Fuerts y Putnam (40) proponen una serie de criterios, obedeciendo la filosofía de los postulados de Koch, las cuales deben de reunirse para aceptar como válida la hipótesis de un fenómeno de interferencia entre plantas, mediadas por agentes químicos o aleloquímicos:
1. Identificación y cuantificación de los síntomas de interacción, excluyendo la posibilidad del fenómeno de competición.
2. Aislar, identificar, ensayar y sintetizar el agente alelopático.
3. Simulación de la interferencia mediante el suministro del agente alelopático en las mismas condiciones observada en la naturaleza.
4. Cuantificación de la dinámica de liberación, movimiento y absorción por organismos receptores del agente alelopático.
Según los datos bibliográficos consultados, no existen antecedentes donde todos y cada uno de estos requerimientos sean satisfechos, sobre todo los referentes a la absorción del aleloquímico por la especie receptora, liberado de manera natural por una planta donadora. La gran mayoría de los estudios llevados a cabo están dirigidos al aislamiento, identificación y síntesis de agentes aleloquímicos, midiendo su actividad mediante bioensayos, estrictamente diseñados y optimizados para probar la hipótesis de la naturaleza de su potencial alelopático (41,42).
En términos generales, estos bioensayos están basados en objetivos bien específicos, siendo dependiente de la naturaleza de la dualidad planta receptora/planta donadora y a las características del agente potencialmente alelopático. Colectivamente, estos objetivos incluyen algunas de las siguientes características del fenómeno de la alelopatía:
· Presencia y liberación de aleloquímicos en la planta denominada donadora.
· Inhibición de los parámetros de crecimiento de la planta denominada receptora e interferencia del aleloquímico con sus procesos fisiológicos.
· Destino final del aleloquímico en el suelo.
· Movimiento del aleloquímico en el suelo, y su absorción por la planta donadora.
· Fenómenos de destoxificación del aleloquímico absorbido por la planta receptora.
· Efecto del aleloquímico sobre la ecología del suelo y dinámica de nutrientes.
· Interacción con promotores, inhibidores y sustancias neutras en el ambiente suelo.
Bajo estas directrices, se han desarrollado una gran diversidad de bioensayos para el estudio de los efectos de los aleloquímicos aislados o liberados por plantas, sobre el crecimiento y distribución poblacional de otras en su vecindad, donde se incluyen los microorganismos (43-46). En otras palabras, la interferencia de estos aleloquímicos sobre las rutas metabólicas o el funcionamiento fisiológico normales de la planta receptora. En esta dirección, la mayoría de los bioensayos se han centrado en la identificación de aleloquímicos potenciales, evaluar sus efectos fitotóxico o su efecto en el crecimientos de plantas, asociadas o no a la productora (plantas receptoras estándares o malas hierbas), con el objetivo de implementar relaciones estructura actividad, que incluyen metabolitos secundarios, aleloquímicos y análogos sintéticos. Todo ello con la filosofía de que puedan ser aplicados como herbicidas, o como aditivos en agentes químicos reguladores del crecimiento (47).
Cada uno de estos bioensayos han sido aplicados de manera exitosa, pero desarrollados y ajustados según diversas variables, como especie receptora, su etapa de desarrollo, soporte del medio nutritivo y el parámetro a medir. Toda esto permitirá realizar las interpretaciones o relaciones entre la actividad (valor del parámetro) y la estructura química del aleloquímico (48-59).
A pesar del amplio espectro de parámetros medidos y controlados en estos bioensayos, su extrapolación a los procesos en el ecosistema natural ha sido tema de discusión (60), y estos debates han originado recientemente el desarrollo de nuevas metodologías para el estudio de la interacción planta donadora-planta receptora (61,62); aportando principalmente una mayor naturalidad al sistema planta-planta del bioensayo.
Sin embargo, en los inicios del desarrollo de estos bioensayos, se cometieron muchos errores metodológicos (63), y aunque sin duda han sido corregidos en los desarrollados durante los últimos 15 años (64), éstos han contribuido a un aumento en la poca credibilidad de lo que pueda representar el fenómeno de la alelopatía en la distribución poblacional de las plantas.
Sobre esto en particular, es curioso observar que, aun siendo la ecología una de las ciencias que más generosa ha sido en cuanto a la aceptación de sus demostraciones experimentales, sin la aclaración minuciosa de los mecanismos microscópicos subyacentes del sistema (21), son los ecólogos los que han mostrado mayor escepticismo en cuanto a la realidad del fenómeno de la alelopatía (65, 66).
Desde su punto de vista, el fenómeno de la alelopatía tiene que ser estudiada en sus fundamentos básicos, principalmente en la determinación de los mecanismos microscópicos subyacentes, entender el modo en que operan, y a partir de esta información, otorgar el peso específico que el fenómeno tiene en la comunidad de las plantas de un ecosistema.
Métodos de estudio y La evidencias del fenómeno alelopático
Sin duda, los mayores avances sobre el fenómeno de la alelopatía se han centrado más en la aplicación que en la esencia del fenómeno, de tal manera que la mayoría de los trabajos encontrados sobre la alelopatía están dirigidos en el aislamiento, la elucidación estructural y la determinación de alguna propiedad o actividad biológica de metabolitos secundarios.
Sin embargo, el estudio de la esencia del fenómeno alelopático ha mostrado menor atención, pues éste resulta un sistema más complejo, y desde el punto de vista ecológico, la principal limitación en su estudio ha sido, como hemos indicado, la factibilidad de separar el efecto alelopático de otros procesos ecológicos (33). No obstante, a pesar de esta dificultad, se han desarrollado algunas metodologías para este fin (18, 22, 67,68). En ellas el carácter multidisciplinar de la alelopatía se ha puesto de manifiesto, y esta característica ha aumentado la dificultad de la demostración del fenómeno. Incluso, algunos sugieren que es imposible separar los efectos alelopáticos de otros procesos que operan simultánea o secuencialmente en la naturaleza (44).
Los indicadores de la presencia del fenómeno no son muchos, y para ser convincente, los estudios se enmarcan en la supresión de la posibilidad de que ocurran otras interacciones. Debido a la complejidad en la separación de un fenómeno de otro en la naturaleza, todo indica que es casi imposible demostrar el fenómeno de la alelopatía (68).
Uno de los resultados que ha demostrado, indicios tanto del carácter multidisciplinario como de la interferencia por el fenómeno alelopático, es el publicado por Bais et al. (69), donde convergen diversas ramas de estudio, como la ecología, la fisiología, la bioquímica y estudios citológicos; en la investigación del desplazamiento de una especie nativa por una invasora exótica, y que éste desplazamiento es atribuido a la acción de (±) catequín como aleloquímico, el cual fue dosificado a las especies receptoras a las mismas concentraciones encontradas en el campo [80-600 mg de catequin.g-1 de suelo seco.]. Estos autores caracterizaron el mecanismo por el cual (-)-catequin suprime la planta nativa, cuyo modo de acción transcurre con la producción de oxígeno activo en la raíz, el cual activa una cascada de señales medidas por la producción de iones Ca(II), donde se inician cambios importantes en la expresión genética y finalmente, la muerte del sistema radicular.
Antecedentes como el descrito han demostrado la interferencia de un compuesto, sobre la comunidad de varias especies de plantas, procedimiento que implica la determinación de varios requisitos descritos anteriormente implementados en la secuencia de la interacción mediada por el fenómeno alelopático, que van desde la identificación del agente alelopático, hasta su interacción, no solo con la planta receptora, sino incluso su interacción con sustancias del medio.
En general, los trabajos referidos al fenómeno alelopático, están centrados en el estudio de la dinámica de la biosíntesis y acumulación del potencial aleloquímico en las plantas (70-74), estudios de la liberación mediante los exudados de la raíz (75,76), de los efectos de la descomposición de su tejido vegetal (77), estabilidad química del aleloquímico una ves traslocado al ambiente (78-80), la medida o registro de alguna actividad biológicas (81-83) y algunos datos sobre su absorción y detoxificación por las potenciales especies receptoras (84-87).
Los diseños experimentales desarrollados para el estudio de cada uno de estos procesos han sido evaluados separándolos de otros posibles fenómenos que puedan ocurrir en el sistema, por ejemplo, medir los niveles de aleloquímicos en los exudados de la raíz, evitando los procesos de biotransformación y de reabsorción del aleloquímico. Aunque estos diseños constituyen estudios sobre un sistema ideal, si se compara con lo que pueda ocurrir en la naturaleza, en conjunto, sus resultados constituyen una información valiosa que han contribuido a entender y ampliar las variables que intervienen en el estudio del fenómeno alelopático.
Más recientemente, se han desarrollado bioensayos para evaluar el potencial alelopático y niveles de exudados de manera simultánea (60,61), donde la planta a la cual se evaluará el potencial alelopático (denominada especie donadora) es mantenida en un cocultivo con una especie (denominada especie receptora) que servirá de modelo para estudiar los efectos causados por la presencia de la especie donadoras. Así como en la mayoría de los trabajos realizados en el campo de la alelopatía, la optimización del diseño experimental es un elemento importante para alcanzar los objetivos planteados, representando una de las variables más importantes en el estudio de las interacciones planta-planta, de cuyos resultados dependerá la interpretación de los resultados, y en gran medida definirá el alcance de los mismos.
Para el diseño del bioensayo de cocultivo; aunque más próximo a una situación real, con el crecimiento de dos especies, con el objetivo de dar un seguimiento a los compuestos involucrados en la interacción, han de considerarse diferentes variables, dependientes de las características de las planta seleccionadas y del agente químico (Figura 1). Características de la especies donadora como velocidad de crecimiento, niveles de aleloquímicos liberados mediante exudado de raíz, densidad de plantas, etc., son variables fundamentales para el registro del aleloquímco en todo el sistema, incluyendo este a la especie receptora.
Por ejemplo, se ha demostrado que en el caso de las benzoxazolinonas, la mayor cantidad de estos compuestos son liberados mediante exudados de la raíz durante sus estados iniciales de crecimiento (61), por lo cual, tiempos de pregerminación muy prolongados, implicarían una menor concentración de los aleloquímicos en el cocultivo como consecuencia del trasplante; permaneciendo una cantidad significativa de compuestos en el medio empleado para la pregerminación. Así, para este tipo de compuestos, es conveniente que el tiempo de pregerminación sea suficiente para asegurar un número predeterminado de plántulas viables, pero que no represente una pérdida considerable de los compuestos en los exudados.
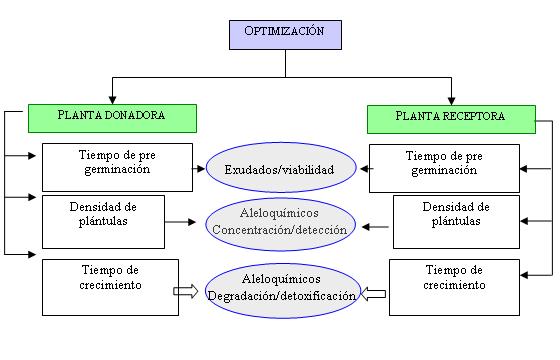
Figura 1. Variables propuestas para la optimización del bioensayo en cocultuivo en el estudio de la interacción mediadas por agentes químicos entre plantas.
Sin embargo, para el caso de las plantas receptoras, el tiempo de pregerminación estará determinado por el nivel de inhibición del crecimiento provocado por el aleloquímico (sensibilidad), de esta manera, si el aleloquímico muestra altos niveles de inhibición de la raíz, por ejemplo, tiempos muy cortos de crecimiento antes de su transplante, podría ocasionar la inviabilidad de las plantas y del bioensayo.
El compromiso en el tiempo de pregerminación dependerá de la dinámica de liberación del compuesto por parte de la planta donadora y los niveles de fitotoxicidad registrados frente a la planta receptora, que permitan logran una máxima sensibilidad en el bioensayo. Este desfase entre el tiempo de crecimiento de ambas plantas, antes de su ensayo de interacción mediante su cocultivo, es un parámetro a considerar en cualquier ensayo.
Por otra parte, la densidad de las plántulas en el bioensayo, y el tiempo de cocultivo, son también parámetros importantes, pues es lo que determinará la posibilidad de detectar el aleloquímico en el tejido de la planta receptora. Así, altas densidades de la planta donadora garantizan una mayor concentración del aleloquímico en el exudado, pero al mismo tiempo, es importante que esta densidad no ocasione una inhibición total de crecimiento de la especie receptora, imposibilitando de esta manera el cocultivo. Por otra parte, períodos largos de cocultivo podría permitir a las plantas receptoras una resistencia hacia el aleloquímico, mediante sus procesos naturales de resistencia, como la detoxificación, disminuyendo de este modo los niveles endógenos, y dificultando su detección. De esta manera, tanto las características de crecimiento, liberación del aleloquímico, como la densidad en el ensayo de la planta donadora como la receptora, han de ser optimizados según las necesidades u objetivos propuestos.
Por ejemplo, para una máxima sensibilidad del ensayo, estas variables han de ser controladas tal que, por ejemplo, la densidad de la planta donadora y el grado de afectación de la receptora, se ajusten a una curva dosis respuesta, de manera de poder calcular parámetros tales como ED50, que representa la densidad de plántula que causa un 50% de reducción del parámetro de crecimiento evaluado respecto a una población control (60). Esta valor de ED50 nos permitirá determinar cual especie donadora presenta mayor actividad alelopática, que parámetro evaluar (peso y/o longitud del tallo o de la raíz), arroje la máxima sensibilidad en el diseño del bioensayo.
Por otro lado, si el objetivo del ensayo es un seguimiento del aleloquímico desde su biosíntesis en la planta donadora hasta su absorción por la receptora, estas variables no se ajustaran a una función dosis-respuesta, sino aquellas condiciones que me permitan la detección del aleloquímico objeto de estudio en todo el sistema. Como consecuencia, los parámetros y/o variables de diseño del bioensayo se complican, pues el método de análisis del aleloquímico formará parte de una variable más considerar, donde las características instrumentales, como el límite de detección y de cuantificación (88, 89), son los que gobernaran las variables biológicas en el diseño del bioensayo (crecimiento y densidad de las plantas).
A pesar de que en un diseño de cocultivo muchas variables pueden ser controladas de tal manera que solo opere la interferencia gobernadas por compuestos químicos, donde competencia por nutrientes, luz, o el efecto de cambios de otros parámetros tales como el pH, sean anuladas; el estudio es extremadamente complicado, pero este diseño representa uno de los que mayormente refleja una condición natural.
El reto que hoy día se presenta es estudiar este sistema con el objetivo de seguir el aleloquímico desde la especie receptora hasta la donadora, de una manera dinámica; es decir, durante el tiempo en que las dos especies estén enfrentadas. El poder determinar toda la ruta del aleloquímico, significaría dar una evidencia contundente de la traslocación de un aleloquímico sintetizado por una especie, liberado al ambiente, detectado en los tejidos de la especie receptora, donde además se registren efectos anormales en sus parámetros de crecimiento. Todo ello representaría una evidencia del fenómeno alelopático, hoy día pendiente de ser demostrado.
El fenómeno alelopatico y la búsqueda de herbicidas.
Durante muchos años, los estudios sobre alelopatía fueron llevados a cabo principalmente por botánicos, dirigiendo sus estudios en función de tres directrices: a) estudiar la práctica del uso del desecho de cosechas en coberturas, b) rotación de cultivos o inter cosechas, y c) estudiar el efecto de extractos de plantas con propiedades alelopáticas en el rendimiento de cosechas.
Más recientemente, debido al mejoramiento y refinamiento de las técnicas de de aislamiento y caracterización, sumado al diseño novedoso de cultivo y bioensayo, los trabajos de investigación están dirigidos a la determinación de la naturaleza estructural de los productos naturales que realmente son responsables de los efectos fitotóxicos observados, dándole un resurgimiento a la alelopatía, convirtiéndola en un “fenómeno diagnóstico” para la búsqueda de moléculas con actividades biológicas importantes. Así, la alelopatía es usada hoy día como una herramienta para identificar especies de plantas que puedan presentar efectos fitotóxicos, las cuales contendrán especies químicas que muestren esta actividad sobre especies de malas hierbas no resistentes y sus biotipos resistentes (Figura 2).
Estos productos que eliminan la competencia de otras especies, o incluso que estimulan la germinación o el crecimiento de otras especies, han despertado cierto interés, principalmente a la posible aplicación de estos en la agricultura, debido, entre otras propiedades, a su actividad herbicida de una manera selectiva (90-92).
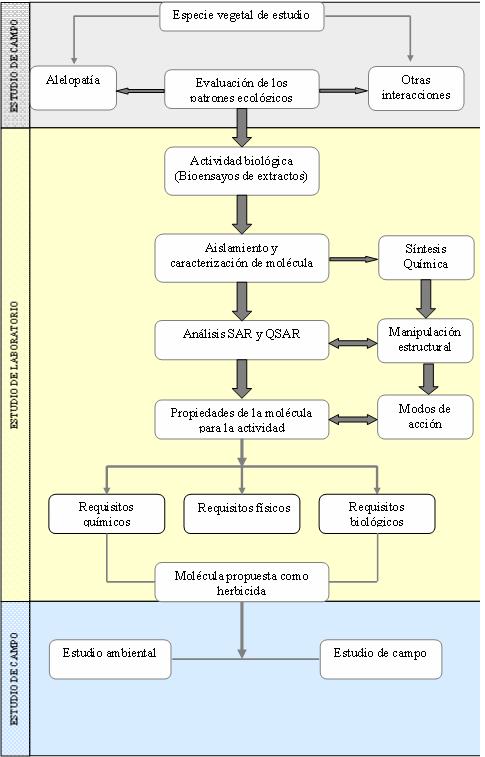
Figura 2. Niveles de estudios abordados en la búsqueda de herbicidas a partir de plantas.
Este hecho a despertado un interés desde el punto de vista agronómico, pues cada ves es más imperante la necesidad de encontrar nuevos herbicidas, entre otras causas, por la proliferación de malezas que han mostrado resistencia frente a los herbicidas convencionales, originándose biotipos muchas más difíciles de controlar (93-96).
Además de la parición de estos biotipos resistentes, la necesidad de generar herbicidas que produzcan un menor impacto en el ambiente, y de cuyos residuos (productos de sus biotransformaciones) sean inocuos para muchos organismos, entre ellos el hombre, ha originado una creciente explotación de las fuentes naturales de compuestos, que en principio, son más específicos en su modos de acción, con productos de biotransformación más tolerables por el ambiente.
Sin embargo, en la actualidad, la alelopatía y los efectos fitotóxicos observados en determinadas moléculas, esta siendo vista no como una fuente directa de compuesto activos, sino como una fuente de estructuras bases, a partir de las cuales, mediante modificaciones sintéticas, darán origen a potenciales herbicidas. Así, una gran diversidad de productos con potencial alelopático han sido encontrados, y de su evaluación fitotóxica, propuestas como moléculas bases para el desarrollo de herbicidas, donde se encuentran terpenos (97), cumarinas (98), benzoquinonas (99) y alcaloides (100) (Figura 3).
A partir de las modificaciones estructurales sintéticas, se originan una serie de derivados modificados, formando una librería de compuestos. La evaluación de las actividades fitotóxicas sobre diferentes especies o series de esta librería, permiten llevar a cabo relaciones estructura actividad (SAR) y estructura reactividad cuantitativas (QSAR) (101-104), que determinen los rasgos estructurales necesarios para la actividad, así como ciertos parámetros (operadores y descriptores moleculares) que indicaran el peso específicos de algunas propiedades fisicoquímicas que controlan las actividades.
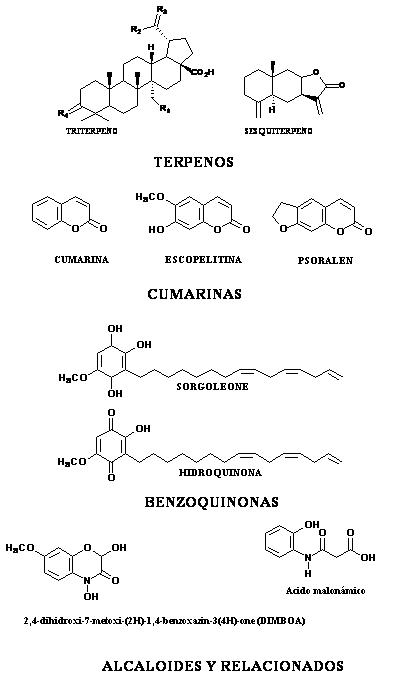
Figura 3. Estructura de algunos metabolitos de origen secundario con propiedades fitotóxicas.
De esta manera es posible manipular sintéticamente la molécula con el objeto de acentuar los rasgos estructurales y físicos a fin de mejorar la actividad bajo estudio, en el caso de herbicidas, sus actividades fitotóxicas. Así, con este tipo de estudio, se han logrado proponer diferentes moléculas con propiedades herbicidas, cuyo origen inicial ha sido el aislamiento de sus estructuras madres a partir de fuentes naturales (Figura 4).
Como un ejemplo de esta metodología, Macías y colaboradores (105) han partido de moléculas de origen natural en la búsqueda de modelos de herbicidas. Tomando como molécula modelo el esqueleto de las benzoxazinonas (Figura 5), una familia de compuestos detectados en los exudados de la raíz de plantas como el maíz, el trigo y el centeno (106-109), los cuales han mostrado una amplia variedad de actividades biológicas, entre ellas las fitotóxicas y alelopáticas (110-116).
Los resultados sobre estudios SAR de las modificaciones sobre las estructuras de las benzoxazinonas (Figura 4, 117), encontraron que la actividad fitotóxica es una función de los efectos estéricos y electrónicos causados por los sustituyentes introducido en el esqueleto base, donde el volumen molecular y momento dipolar resultan los más relevantes. A partir de los estudios QSAR, se encontró que la liposolubilidad (la solubilidad en medio oleoso) es un factor determinante en la actividad fitotóxicos, y que en algunos casos, los derivados muestran una mayor actividad fitotóxica y selectividad sobre la especie receptora que sus productos naturales de partida (105).
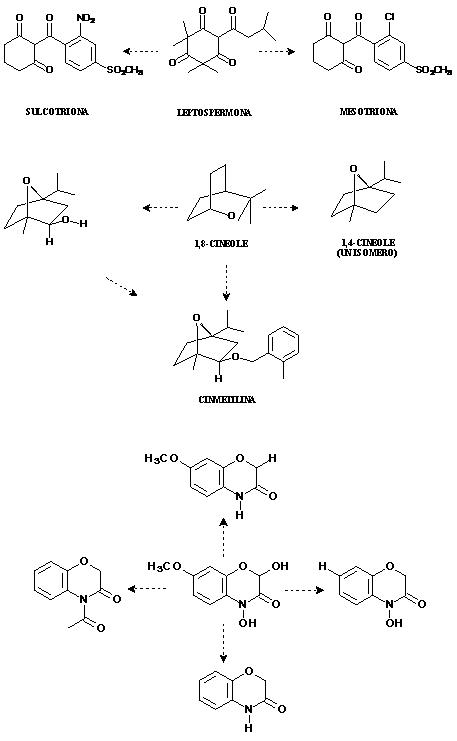
Figura 4. Modificaciones sintéticas sobre moléculas de origen natural para la búsqueda de modelos de herbicidas.
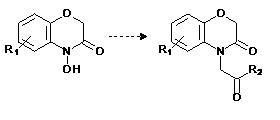
Figura 5. Estructura general de los derivados de las benzoxazinonas en los estudios SAR y QSAR para la búsqueda de modelos de herbicidas naturales.
Productos naturales explotados como herbicidas comerciales.
La obtención de herbicidas y agroquímicos basados en fuentes naturales son atractivos por varias razones. Primero, la mayoría de ellos son solubles en agua, y como resultado, es posible que sean activos a muy bajas concentraciones, reduciendo su presencia en el ambiente. Son más inocuos al ambiente, además de presentar modos de acción no registrados a los herbicidas hoy día existente, convirtiéndose en una promesa en el control de los biotipos resistentes.
Sin embargo, el desarrollo del uso de herbicidas fuentes naturales en la industria agroquímica ha sido muy limitado. Varias razones o variables han de ser consideradas para llegar a explotar este aleloquímico como agente de control de malas hierbas, donde es posible citar: a) Cantidad de sustancia activa aislada, generalmente muy limitada para los diferentes tipos de bioensayos. b) Necesidades de pruebas toxicológicas y ambientales. El hecho de que el producto sea de origen natural, no garantiza que sea seguro al ambiente, y de hecho, la mayoría de los tóxicos para los mamíferos son de origen natural, y dentro de estos, muchos provienen de las plantas. c) Originar compuestos derivados a partir del natural, mediante manipulación de la estructura molecular o síntesis química. Una de las características en general de los productos naturales es su extremada complejidad estructural, lo cual es un factor limitante en la obtención de series que permitan realizar estudios SAR y QSAR, d) una ves detectado el producto activo, o su análogo modificado, es necesario la extracción en masa del compuesto de su fuente natural, o en su defecto, una ruta sintética relativamente sencillas y rentable.
Para esto, la especie o variedades deben de producir grandes cantidades del producto con propiedades herbicidas, donde la manipulación genética para incrementar la producción del compuesto se convierte en una variable y factor limitante en el proceso de desarrollo de herbicidas. Incluso, muchos herbicidas a bajas dosis pueden causar un incremento en los niveles de fitoalexinas presentes en muchas plantas de interés agronómico, con lo cual el efecto de este producto sobre la química de las plantas a proteger debe de estar incluido en el protocolo de desarrollo de bioherbicidas. e) Por último, recientemente el desarrollo de herbicidas a partir de fuente natural se ha visto limitada por la patentabilidad de los compuestos o moléculas. La patente puede ser efectuada tanto sobre el compuesto natural así como compuestos sintéticos, hecho este que limita de manera decisiva los estudios de actividad biológica.
A pesar de estas variables que dificultan la búsqueda de compuestos modelos de herbicidas, existen casos donde moléculas de origen natural están siendo explotadas como herbicidas comerciales, tales Cinmetilina, Bialafos, Glufosinatos y Dicamba.
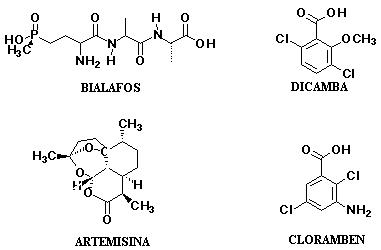
Figura 6. Compuestos aislados a partir de plantas que han sido explotados como herbicidas comerciales.
Cinmetilina ha sido aplicado para el control de malas hierbas durante periodos corto de tiempo (118). Bialafos es actualmente comercializado en Japón bajo el nombre de Herbace® (119). Un derivado químico de la leptospermone, un componente de los aceites esenciales de Leptospermum scoparium (120), origino al mesotrione (121), el ingrediente activo del herbicida comercial Callisto ®, un herbicida sistémicos, selectivo y de aplicación post emergente, el cual puede controlar malas hiervas que han desarrollado resistencia a la antrazina, principalmente aplicado en cultivos de maíz. Otro derivado de la leptospermona, la sulcortiona, ha sido comercializado en Europa por la Bayer CropScience bajo el nombre de Mikado®, siendo también un herbicida post emergente, utilizado ampliamente en los cultivos del maíz y caña de azúcar frente a una gran variedad de malas hierbas.
Estos ejemplos de productos naturales con una amplia aplicación en el control de malas hiervas, demuestra la aplicabilidad de la explotación de los productos de origen natural como fuente de una amplia variedad de compuestos con actividades biológicas importantes. Aquí solo se ejemplifican aquellos compuestos de origen vegetal para el control de malas hierbas, sin embargo, otras fuentes naturales como metabolitos a partir de cultivo de microorganismos, hongos, microbios, etc., son también una fuente rica en compuestos novedosos, no solo con actividad fitotóxica, sino insecticidas (122,123), repelentes (124-127) y larvicidas (128-131), entre otras actividades.
Conclusiones
La interacción entre plantas, promovidas por agentes químicos, ha sido un tema de extensa investigación, donde la ecología y su variante, la química ecológica, ha sido la encargada de estudiar y explotar este fenómeno. Aunque la demostración del fenómeno, resumido en la detección del compuesto liberado por una especie, en el tejido vivo de otra especie, es una tarea pendiente en el estudio de los principios básicos de la acción alelopática, es sin duda las aplicaciones que el fenómeno ha permitido lo que le ha dado una mayor importancia.
Desde sus inicios hasta nuestros días, la alelopatía ha experimentado un desarrollo significativo, encontrándose que es un mecanismo de interacción muy complejo, confiriéndole un carácter interdisciplinario. De esta manera, encontrar su explotación, como por ejemplo, la búsqueda de herbicidas naturales, requiere de la participación de botánicos, ecólogos, químicos orgánicos y analíticos, e inclusos biólogos moleculares.
A pesar de esta convergencia de ciencias, el estudio de la búsqueda de herbicidas naturales no es una tarea fácil, donde muchas limitaciones son encontradas. Por ejemplo, el alto costo de su obtención a partir de la fuente natural, moléculas con rutas de síntesis complejas, una actividad muy restringida y sobre todo, los efectos fitotóxicos que puedan mostrar sobre organismo no receptores.
Otros elementos ecológicos recientemente han sido considerados en el estudio del fenómeno dirigido a la búsqueda de modelos de herbicidas, tales como factores edáficos, hábitat y clima, han sido incluidas como variables que determinaran el destino, persistencia, y disponibilidad, tanto desde un punto de vista cuantitativo como cualitativo, de los aleloquímicos liberados en la rizósfera.
Estas variables determinaran la especie verdaderamente activa y los niveles de concentración a las cuales ellos muestran los efectos observados, dos factores fundamentales para el desarrollo de compuestos altamente fitotóxicos. De esta manera, la estabilidad en el suelo ha sido incluida en los estudios de los compuestos exudados por la raíz, donde es necesario determinar su persistencia en suelo, su absorción física y química con las partículas del suelo y, de mayor importancia, las diferentes especies a la que es convertido por los procesos biológicos de la rizósfera.
Por último, son los químicos orgánicos sintéticos los que tendrán la última palabra en el desarrollo de bioherebicidas, pues la síntesis de la amplia variedad de productos naturales con actividad alelopática, así como de modificaciones estructurales sistemáticas y controladas que generen estructuras análogas, son fundamentales para un estudio más detallado de las necesidades estructurales para la actividad observada; es decir, llevar a cabo relaciones SAR y QUSAR, y en última instancia, determinar los modos de acción de los agentes naturales y sus análogos sintéticos.
Referencias
1. Coley P. 1983. Herbivory and defensive characteristics of tree species in a lowland tropical forest Ecol. Monog. 53:209-233.
2. Aide T. 1993. Patterns of leaf development and herbivory in a tropical understudy community Ecology 74:455-466
3. Rosenthal G, Berembaum M. 1991. Herbivores, Their Interactions with Secondary Plant Metabolites. 2nd ed. Vol. 1. San Diego/CA: Academic Press
4. Tallamy D, Raupp M. 1991 Phytochemicals Induction by Herbivores, New York: John Wiley& Sons.
5. Singh H, Batish D, Kohli R. 2003. Allelopathic Interactions and Allelochemicals: New Possibilities for Sustainable Weed Management Crit. Rev. Plant Sci. 22:239-311.
6. Welden, C., Slauson W. 1986. The intensity of competition verses its importance: an overlooked distinction and some implications Q Rev Bot 61:23–44
7. Callaway R, Nadkarni N, Mahall B. 1991. Facilitating and interfering effects of Quercus douglasi in central California Ecology 72:1484–1499
8. Chapin F., Walker L, Fastie C, Sharman L. 1994. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska Ecol. Monogr 64:149–175
9. Muller C, Hanawalt R, McPherson J (1969) Allelopathic control of herb growth in the fire cycle of California chaparral Bull Torrey Bot Club 95:225–231
10. Rice E. 1974. Allelopathy. Academic Press, New York.
11. Connell J. 1983. On the prevalence and relative importance of interspecific competition: evidence from field experiments Am Nat 122:661- 696
12. Schoener T. 1983. Field experiments on interspecific competition Am Nat 122:240–285
13. Caldwell M, Eissenstat D, Richards J, Allen M. 1985. Competition for phosphorous: differential uptake from dualisotope-labeled interspaces between shrubs and grass Science 229:384–384
14. Caldwell M, Richards J, Manwaring J, Eissenstat D. 1987. Rapid shifts in phosphate acquisition show direct competition between neighboring plants Nature 327:615–616
15. Mahall B.; Callaway R. 1991. Root communication among desert shrubs Ecology 88:874–876
16. Mahall B, Callaway R. 1992. Root communication mechanisms and intracommunity distributions of two Mojave Desert shrubs Ecology 73:2145–2151
17. Fuerst E, Putman A. 1983. Separating the competitive and allelopathic components of interference: theoretical principles J. Chem Ecol 9:937–944
18. Thijs H, Shann J, Weidenhamer J. 1994. The effect of phytotoxins on competitive outcome in a model system Ecology 75: 1959–1964.
19. Harper J. 1977. Population biology of plants. Academic Press, London.
20. Connell J. 1990. Apparent versus “real” competition in plants. In: Grace JB, Tilman D (eds) Perspectives on plant competition. Academic Press, San Diego, Calif., pp 9–26
21. Williamson G. 1990. Allelopathy, Koch’s postulates, and the neck riddle. In: Grace J, Tilman D. eds. Perspectives on plant competition. Academic Press, San Diego, Calif., pp 143–162.
22. Nilsson M. 1994. Separation of allelopathy and resource competition by the boreal dwarf shrub Empetrum hermaphroditum Oecologia 98:1–7.
23. Ridenour W, Callaway R. 2001. The relative importance of allelopathy in interference: the effects of an invasive weed on a native bunchgrass Oecologia 126:444–450
24. Rice E. 1984. Allelopathy, 2da ed. Academic Press, Orlando, FL.
25. International Allelopathuy Society (1996) Constitutions.Drawn up duraing First World Congress on Allelopathy: A Science for the Future. Cadiz, Spain. http://www-ias.uca.es/bylaws.htm#CONSTI. Acceso en marzo 2008.
26. Inderji, Dakschini K. 1994. Algal allelopathy Bot. Rev. 60:182-196
27. Wardle D, Nilsson M, Gallet C, Zackrisson O. 1998. An ecosystem-level perspective of allelopathy Biolo. Rev. 73:305-319.
28. Inderjit, Callaway R. 2003. Experimental designs for the study of allelopathy Plant Soil 256:1-11.
29. Suikkanen S, Fistarol G, Graneli E. 2004. Allelopathic effects of the Baltic cianobacteria Nodularia spumigena, Aphanizomenon flosaquae and Anabaena lemmermanni on algal monocultures J. Exp. Marine Biol. Ecol. 308: 85-11.
30. Fitter A. 2003. Making allelopathy respectable Science 301:1337-1338.
31. Hierro J, Callaway R. 2003. Alleoptahy and exotic plant invasión Plant Soil 256:29-39.
32. Vivanco J, Baiz H, Stermitz T.,Thelen G, Callaway R. 2004. Biogeochemical variation in community responses to root allelochemistry: novel weapons and exotic invasion Ecol. Lett. 7:285-292.
33. Reigosa M, Sánchez-Moreiras A, González L. 1999. Ecophysiological Approach in Allelopathy Crit. Rev. Plant Sci. 18:577-608.
34. Chiapusio G, Sánchez A, Reigosa M, González L, Pellissier F. 1997. Does the use of germination indexes reflect allelochemicals effects on germination processes? J. Chem. Ecol. 21: 1400–1411.
35. Pickering S. 1917. The effect of the one another Ann. Bota. 31: 181-187.
36. Pickering S. 1919. The action of crop on another J. Roy. Hort. Soc. 43:372-380.
37. Citado en: Rice E. 1979. Allelopathy - an update Bot. Rev. 45:15-109.
38. Traducción de: Morton A. 1981. History of botanical science. Academic Press, London.
39. Koch R. 1884. The etiology of tuberculosis [Koch’s postulates.] The Germ. Theory Dis.116-118. Koch R. 1884. Die Aetiologie der Tuberkulose.Mittbeihgen aus dem Kaiserlichen Gesundbeitsamte,Vol. 2, pages 1-88.
40. Fuerst E, Putnam A. 1983. Separating the competitive and allelopathic components of interference: theoretical principles J.Chem. Ecol. 9:937-44.
41. Wolfson J. 1988. Bioassay techniques: an ecological perspective J. Chem. Ecol. 14:1951–1963.
42. Inderjit; Nilsen E. 2003. Bioassays and Field Studies for Allelopathy in Terrestrial Plants: Progress and Problems Crit. Rev. Plant Sci. 22:221-238.
43. Inderjit, Del Moral R. 1997, Is separating resource competition from allelopathy realistic? Bot. Rev. 63:221-230.
44. Torquebiau E. 1994. Ecological interactions in agroforestry Personal communication.
45. Vandermeer H. 1980. The ecology of intercropping. Cambridge Univ. Press. Cambridge, U.K
46. Hoagland R, Williams R. 2004. Bioassay – Useful Tools for the Study of Allelopathy. In Allelopathy. Chemistry and Mode of Action of Allelochemicals. Francisco A. Macías, Juan C.G. Galindo, José M.G. Molinillo and Horace G. Cutler eds. CRC Press.
47. Fay P, Duke W. 1977. An assessment of allelopathic potential in Avena germ plasm Weed Sci. 25:224-228.
48. Fujii Y. 2001. Screening and future exploitation of allelopathic plant as alternative herbicides with special referent to hairy vethc J. Crop. Prod. 4:257-275.
49. Fujii Y, Yasuda T. 1990. Methods for screening allelopathic activities by using the logistic function (Richards’ function) fitted to lettuce seed germination and growth curves Weed Res. 35:353-361.
50. Hashem A., Adkins S. 1998. Allelopathic effect of Triticum speltoides on two important weeds of wheat. Plant Prot. Q. 13:33-35.
51. Navarez D, Olofsdotter M. 1996. Relay seeding technique for screening allelopathic rice (Oryza sativa), pp.1175-1181, in Proceedings of the 2nd International Weed Control Congress, Copenhagen.
52. Putnam A, Duke W. 1974. Biological suppression of weeds: Evidence for allelopathy in accessions of cucumber Science 185:370-371.
53. Rimando A, Olofsdotter M, Duker S. 2001. Searching for rice allelochemicals: An example of bioassay-guided isolations Agr. J. 93:16-20.
54. Wu H, Pratley J, Lemerle D, Haig T. 2000. Laboratory screening for allelopathic potential of wheat (Triticum aestivum) accessions again annual ryegrass Aust. Agric. Res. 51:259-266.
55. Wu H, Pratley J, Lemerle D, Haig T, An M. 2001. Screening methods for the evaluation of crop allelopathic potential Bot. Rev. 67:403-415.
56. Devi S, Pellissier F, Prasad M. 1996. Allelochemicals. In: Plant Ecophysiology. Prasad., M. Ed., John Wiley and Sons, New York, 253–303.
57. Inderjit; Dakshini K. 1995. On laboratory bioas-says in allelopathy Bot. Rev. 61:28-44.
58. Saxena A, Singh D, Joshi N. 1996. Allelopathy in agroecosystems Field Crop Abstract 49: 891–900.
59. Inderjit; Weston L. 2000. Are laboratory bioassays for allelopathy suitable for prediction of field responses? J. Chem. Ecol. 26:2111-2118.
60. Belz R, Hurle K. 2004. A Novel Laboratory Screening Bioassay for Crop Seedling Allelopathy J. Chem. Ecol. 30:175-198.
61. Belz R, Hurle K. 2005. Differential Exudation of Two Benzoxazinoids - One of the Determining Factors for Seedling Allelopathy of Triticeae Species, J. Agrc. Food Chem. 53:250-261.
62. Davis L. 1996. Allelopathy tested again Am. Biol. Teacher 58: 263.
63. Ver por ejemplo: Stowe L. 1979. Allelopathy and its influence on the distribution of plants in an Illinois oldfield J. Ecol. 67:1065–1085.
64. Fisher N, Williamson G, Weidenhamer J, Richardson D. 1994. In search of allelopathy in the Florida scrub: the role of terpenoids J. Chem. Ecol. 20:1355–1380.
65. Heldt H. 1997. Plant Biochemistry and Molecular Biology.Oxford University Press, Oxford.
66. Weidenhamer J, Hartnett D, Romeo J. 1989. Densitydependent phytotoxicity: distinguishing resourcecompetition and allelopathic interference in plants J. Appl. Ecol. 26:613-624.
67. Inderjit; Dakshini K. 1988. Allelopathic interference of chickweed, Stellaria media with seedling growth of wheat (Triticum aestivum) Can. J. Bot. 76: 1317–1321.
68. Harper J. 1975. Allelopathy Quarterly Rev. Biol. 50:493-495.
69. Bais H, Vepachedu R, Gilroy S, Callaway R, Vivanco J. 2003. Allelopathy and Exotic Plant Invasion: From Molecules and Genes to Species Interactions Science 301:1377-1380. en: www.sciencemag.org/cgi/content/full/301/5638/1377/DC1
70. Copaja S, Nicol D, Wratten S. 1998. Accumulation of hydroxamic acids during wheat germination. Phytochemistry 50:17-24.
71. Collantes H, Gianoli E, Niemeyer H. 1999. Defoliation affects chemical defenses in all plant parts of rye seedlings J. Chem. Ecol. 25:491-499.
72. Rice C, Park Y, Adam F, Abdul-Baki A, Teasdale J. 2005. Hydroxamic Acid Content and Toxicity of Rye at Selected Growth Stages J. Chem. Ecol. 31:1887-1905.
73. Rice C, Park Y, Adam F, Abdul-Baki A, Teasdale J. 2005. Hydroxamic acid content and toxicity of rye at selected growth stages J. Chem. Ecol. 31:1887-1905.
74. Stochmal A, Kus J, Martyniuk S, Oleszek W. 2006. Concentration of benzoxazinoids in roots of field-grown wheat (Triticum aestivum L.) varieties J. Agric. Food Chem. 54:1016-1022.
75. Huang Z, Terry H, Wu H, An M, Pratley J. 2003. Correlation between phytotoxicity on annual ryegrass (Lolium rigidum) and production dynamics of allelochemicals within root exudates of an allelopathic wheat J. Chem. Ecol. 29:2263-2279.
76. Pérez F, Ormeño-Núñez J. 1991. Difference in hydroxamic acid content in roots and root exudates of wheat (Triticum aestivum L.) and rye (Secale cereale L.): possible role in allelopathy J. Chem. Ecol. 17: 1037-1043.
77. Mathiassen S, Kudsk P, Mogensen B. 2006. Herbicidal Effects of Soil-Incorporated Wheat J. Agric. Food Chem. 54:1058-1063.
78. Zikmundova M, Drandarov K, Bigler L, Hesse M, Werner C. 2002. Biotransformation of 2-benzoxazolinone and 2-hydroxy-1,4-benzoxazin-3-one by endophytic fungi isolated from Aphelandra tetragona Appl. Environ. Microbiol. 68:4863-4870.
79. Fomsgaard I, Mortensen A, Carlsen S. 2004. Microbial transformation products of benzoxazolinone and benzoxazinone allelochemicals-a review Chemosphere 54:1025-1038.
80. Macias F, Oliveros-Bastidas A, Marin D, Castellano D, Simonet A, Molinillo J. 2005. Degradation Studies on Benzoxazinoids. Soil Degradation Dynamics of (2R)-2-O-β-D-Glucopyranosyl-4-hydroxy-(2H)-1,4-benzoxazin-3(4H)-one (DIBOA-Glc) and Its Degradation Products, Phytotoxic Allelochemicals from Gramineae J. Agric. Food Chem. 53:554-561.
81. Sanchez-Moreiras A, Reigosa M. 2005. Whole Plant Response of Lettuce after Root Exposure to BOA (2(3H)-Benzoxazolinone) J. Chem. Ecol. 31: 2689-2703.
82. Macias F, Marin D, Oliveros-Bastidas A, Castellano D, Simonet A, Molinillo J. 2006. Structure-activity relationship (SAR) studies of benzoxazinones, their degradation products, and analogues. Phytotoxicity on problematic weeds Avena fatua L. and Lolium rigidum Gaud J. Agric. Food Chem. 54: 1040-1048.
83. Martyniuk S, Stochmal A, Macias F, Marin D, Oleszek W. 2006. Effects of some benzoxazinoids on in vitro growth of Cephalosporium gramineum and other fungi pathogenic to cereals and on Cephalosporium stripe of winter wheat J. Agric. Food Chem. 54:1036-1039.
84. Schulz M, Wieland I. 1999. Variation in metabolism of BOA among species in various field communities Biochemical evidence for co-evolutionary processes in plant communities? Chemoecology. 9:133-141.
85. Chiapusio G, Pellissier F, Gallet Ch. 2004. Uptake and translocation of phytochemical 2-benzoxazolinone (BOA) in radish seeds and seedlings J. Exp. Bot. 55:1587-1592.
86. Baerson S, Sanchez-Moreiras A, Pedrol-Bonjoch N, Schulz M, Kagan I, Agarwal A, Reigosa M, Duke S. 2005. Detoxification and Transcriptome Response in Arabidopsis Seedlings Exposed to the Allelochemical Benzoxazolin-2(3H)-one. J. Biol. Chem. 280:21867-21881.
87. Hofmann D, Knop M, Hao H, Hennig L, Sicker D, Schulz M. 2006. Glucosides from MBOA and BOA detoxification by Zea mays and Portulaca oleracea J. Nat. Prod. 69:34-7.
88. Wolfender J, Ndjoko K, Hostettmann, K. 2003. L iquid chromatography with ultraviolet absorbance–mass spectrometric detection and with nuclear magnetic resonancespectroscopy: a powerful combination for the on-line structural investigation of plant metabolites. J.Chromatogr. A, 1000, 437–455.
89. Eljarrat E, Guillamon M, Seuma J, Mogensen B, Fomsgaard I, Oliveros-Bastidas A.; Macias F, Stochmal A, Oleszek W, Shakaliene O, Barcelo D. 2004. First European interlaboratory study of the analysis of benzoxazinone derivatives in plants by liquid chromatography J. Chromatogr. A 1047, 69-76.
90. Duke S, Dayan F, Romagni J, Rimando A. 2000. Natural products as sources of herbicides: current status and future trends. Weed Res. 40:99-111.
91. Stonar R, Miller-Wideman M. 1995 Herbicides and Plant growth regulators. In: Agrochemicals from Natural Product., Godfrey, C. Ed., Marcel Dekkert: New York, cap.6, pp.285-310.
92. Fischer N, Weidenhamer J, Riopel J, Quijano L, Menelaou M. 1990. Stimulation of witchweed germination by sesquiterpene lactones: a structure-activity study. Phytochemistry. 29:2479-2483.
93. Heap, I. 1997. The occurrence of herbicide-resistant weed world-wide. Pestic. SCI. 51:235-243.
94. Itoh K, Wang G, Ohba S. 1999. Sulfonylurea resistence in Lindernia microntha, an annual paddy weeds in japan. Weed Res. 39:413-423.
95. Heap I. 2003. The international survey of herbicide resistant weeds. Internet. Febrero 28, 2008. Available online at: www.weedscience.com. Acceso marzo de 2008
96. Sandermann H. 2006. Plant biotechnology biotechnology: ecological case studies on herbicide resistance Trends in Plant Sci. 11, 7: 324-328.
97. Elakovich S, In Biologically Active Natural products: Potential Use in Agriculture. Cutler, H. Ed. ACS Symposium Series 380, American Chemical Society: Washington, DC. 1988.
98. Zobel A. In Principes and Practices in Plant Ecology: Allelochemical Interactions. Inderjit, Dakshini, K.; Foy, C. Eds.; CRC: Boca Raton, Florida. 1999.
99. Netzly D, Riople J, Eljeta G, Butler L. 1988. Germination stimulants of witchweed (Striga asiatica) from hydrophobic root exudates of sorghum (Sorghum bicolor) Weed Sci. 36:441-446.
100. Macías F, Marín D, Oliveros-Bastidas A, Castellano D, Simonet A, Molinillo J. 2005. Structure-Activity Relationships (SAR) Studies of Benzoxazinones, Their Degradation Products and Analogues. Phytotoxicity on Standard Target Species (STS) J. Agric. Food Chem. 53: 538-548.
101. Vyvyan J. 2002. Allelochemical as leads for new herbicides and agrochemicals Tetrahedron. 58:1631-1646.
102. Seydel J. QSAR and Strategies in the Design of Bioactive Compounds. 1985, VCH, Weinheim.
103. Patani G, LaVoie E. 1996. Bioisosterism: A Rational Approach in Drug Design. Chem. Rev. 96: 3147-3176.
104. Duchowicz P, Mercader A, Fernández F, Castro E. 2008. Prediction of aqueous toxicity for heterogeneous phenol derivatives by QSAR. Chemometrics and Intelligent Laboratory Systems 90: 97–107.
105. Macías F, Marín D, Oliveros-Bastidas A, Molinillo J. (2006) Optimization of Benzoxazinones as Natural Herbicide Models by Lipophilicity Enhancement. J. Agric. Food Chem. 54: 9357-9365.
106. Pérez F, Ormeño-Núnez J. 1991. Difference in hydroxamic acid content in roots and root exudates of wheat (Triticum aestivum L.) and rye (Secale cereale L.): Possible role in allelopathy. J. Chem. Ecol. 17: 1037-1043.
107. Pérez F, Ormeño-Nuñez J. 1993. Weed growth interference from temperate cereals: the effect of a hydroxamic-acid-exuding rye (Secale cereale L.) cultivar. Weed Res. 33: 115-119.
108. Wu H, Pratley J, Lemerle D, Haig T. 2001. Allelopathy in wheat (Triticum aestivum). Ann. Appl. Biol. 139:1-9.
109. Niemeyer H, Copaja S, Barria B, 1992. The Triticeae as sources of hydroxamic acids, secondary metabolites in wheat conferring resistance against aphids. Hereditas 116:295-299.
110. Honkanen E, Virtanen A. 1960. Synthesis of some 1,4-benzoxazine derivatives and their antimicrobial activity. Acta Chem. Scand. 14:1214-1217.
111. Escobar C, Sicker D, Niemeyer H. 1999. Evaluation of DIMBOA analogs as antifeedants and antibiotics towards the aphid Sitobion aVenae in artificial diets. J. Chem. Ecol. 25:1543-1554.
112. Schulz M, Friebe A, Kueck P, Seipel M, Schnabl H. 1994. Allelopathic effects of living quack grass (Agropyron repens L.). Identification of inhibitory allelochemicals exuded from rhizome borne roots. Angew. Bot. 68:195-200.
113. Wolf, R. B.; Spencer, G. F.; Plattner, R. 1985. Benzoxazolinone, 2,4-dihydroxy-1,4-benzoxazin-3-one, and its glucoside from Acanthus mollis seeds inhibit velvetleaf germination and growth. J. Nat. Prod. 48:59-63.
114. Macías F., Marín D, Oliveros-Bastidas A, Castellano D, Simonet A, Molinillo J. 2005. Structure-activity relationships (SAR) studies of benzoxazinoids, their degradation products and analogues. Phytotoxicity on standard target species (STS). J. Agric. Food Chem. 53:538-548.
115. Macías F, Chinchilla N, Varela R, Oliveros-Bastidas A, Marín D, Molinillo J. 2005. Structure-activity relationship studies of benzoxazinones and related compounds. Phytotoxicity on Echinochloa crus-galli (L.) P. Beauv. J. Agric. Food Chem. 53: 4373-4380.
116. Macías F, Marín D, Oliveros-Bastidas A, Castellano D, Simonet A, Molinillo J. 2006. Structure-activity relationship (SAR) studies of benzoxazinones, their degradation products, and analogues. Phytotoxicity on problematic weeds Avena fatua L. and Lolium rigidum Gaud. J. Agric. Food Chem. 54:1040-1048.
117. Macías F, Siqueira J, Chinchilla N, Marín D, Varela R, Molinillo J. 2006. New Herbicide Models from Benzoxazinones: Aromatic Ring Functionalization Effects J. Agric. Food. Chem. 54:9843-9851.
118. Bhowmik P. (1988) Cinmethylin for weed control in soybeans. Glycine max. Weed Res. 36:678-682.
119. Klink J., Brophy J, Perry N, Weavers R. 1999. Weavers, β-Triketones from Myrtaceae: Isoleptospermone from Leptospermum scoparium and papuanone from Corymbia dallachiana. J. Nat. Prod. 62: 487-489.
120. Saxena S, Pandey A. 2001. Microbial metabolites as eco-friendly agrochemicals for the next millennium. Appl. Microbiol. Biotechnol. 55: 395-403.
121. Mitchell G, Bartlett D, Fraser T, Hawkes T, Holt D, Townson J, Wichert R. 2001. Mesotrione: a new selective herbicide for use in maize. Pest Manag. Sci. 57:120-128.
122. Isman M, Akhtar Y. 2007. Plant Natural Products as a Source for Developing Environmentally Acceptable Insecticides, in Insecticides Design Using Advanced Technologies Springer Berlin Heidelberg, pp. 235-248
123. Sparks T, Crouse G, Durst G. 2001. Natural products as insecticides: the biology, biochemistry and quantitative structure-activity relationships of spinosyns and spinosoids. Pest Manag. Sci. 57: 896-905.
124. Ansari M, Razdan R. 1985. Relative efficacy of various oils in repelling mosquitoes. Indian J. Malariol. 32:104–111.
125. Barnard D, Xue R. 2004. Laboratory evaluation of mosquito repellents against Aedes albopictus, Culex nigripalpus, and Ochlerotatus triseriatus (Diptera: Culicidae). J. Med. Entomol. 41:726-730.
126. Deshmukh P, Renapurkar D. 1989. Insect repellent activity of Acorus calamus (Linn) against mosquito Culex pipiens fatigans. Pestology. 13:11–14.
127. Govere J, Durrheim D, Baker L, Hunt R, Coetzee M. 2000. Efficacy of three insect repellents against the malaria vector Anopheles arabiensis. Med. Vet. Entomol. 14: 441–444.
128. Kumar A., Dutta G. 1987. Indigenous plant oils as larvicidal agents against Anopheles stephensi mosquitoes. Curr. Sci. 56: 959–960.
129. Perich M, Wells C, Bertsch W, Tredway K. 1994. Toxicity of extracts from three Tagetes against adults and larvae of yellow fever mosquito and Anopheles stephensi (Diptera: Culicidae). J. Med. Entomol. 31:833–837.
130. Saxena R, Harshan V, Saxena A, Sukumaran P, Sharma M, Kumar M. 1993. Larvicidal and chemosterilant activity of Annona squamosa alkaloids against Anopheles stephensi. J. Am. Mosq. Control Assoc. 9: 84–87.
131. Knioa K, Ustab J, Daghera S, Zournajiana H, Kreydiyyeha S. 2008 Larvicidal activity of essential oils extracted from commonly used herbs in Lebanon against the seaside mosquito, Ochlerotatus caspius. Bioresource Technology. 99:763-768.
ISSN 1666-7948
www.quimicaviva.qb.fcen.uba.arRevista QuímicaViva
Número 1, año 7, Abril 2008
quimicaviva@qb.fcen.uba.ar