Toxicocinética del metilazinfos e inhibición in vitro de la actividad colinesterásica por metilazinfos-oxón en los invertebrados Biomphalaria glabrata y Lumbriculus variegatus
Luis Claudio Cacciatore, Noemí Rosario Verrengia Guerrero,
Adriana Cristina Cochón*
Departamento de Química Biológica, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, Nuñez, 1428, Buenos Aires, Argentina.
E-mail: adcris@qb.fcen.uba.ar
Recibido 25 de Octubre 2013- Aceptado 29 de Octubre 2013
Resumen
Los pesticidas organofosforados ejercen su toxicidad por inhibición de la
enzima acetilcolinesterasa, causando neurotoxicidad. La presencia de esta
enzima en invertebrados y vertebrados confiere a esta clase de pesticidas
una enorme toxicidad hacia blancos no deseados. El metilazinfos es un
insecticida organofosforado que debe sufrir una activación metabólica a
metilazinfos-oxón para poder inhibir la actividad colinesterasa. En trabajos
anteriores se había demostrado que el oligoqueto Lumbriculus variegatus es
mucho más sensible a la exposición in vivo a metilazinfos que el gasterópodo
Biomphalaria glabrata. En el presente trabajo se investigó si esa
sensibilidad diferencial se debía a diferencias en las cinéticas de
absorción y/o eliminación del pesticida o a diferencias en la sensibilidad
in vitro de las colinesterasas de ambos invertebrados al oxón derivado.
Utilizando un modelo unicompartimental de primer orden se pudo comprobar que
la absorción y eliminación del pesticida son muy rápidas en ambas especies
con factores de bioconcentración muy similares. Por el contrario, se
necesitó una concentración 1500 veces mayor de metilazinfos-oxón para
inhibir un 50% la actividad colinesterasa en los extractos del gasterópodo
que en el oligoqueto. Estos resultados estarían indicando que la diferente
sensibilidad de ambas especies a la inhibición de la actividad colinesterasa
por metilazinfos se debería más a factores toxicodinámicos que a factores
toxicocinéticos.
Palabras clave: metilazinfos, invertebrados, toxicocinética, colinesterasa
Toxicokinetics of azinphos-methyl
and in vitro inhibition of cholinesterase activity by azinphos-methyl-oxon
in two freshwater invertebrates: Biomphalaria glabrata and Lumbriculus
variegatus
Abstract
Organophosphate pesticide toxicity occurs through inhibition of the enzyme
acetylcholinesterase, resulting in neurotoxicity. The presence of this
enzyme in invertebrates and vertebrates confers to this class of pesticides
a great toxicity to non target organisms. Azinphos-methyl is an
organophosphate insecticide that must undergo metabolic activation to
azinphos-methyl-oxon in order to inhibit cholinesterase activity. In a
previous study, a greater sensitivity of Lumbriculus variegatus when
comparing with Biomphalaria glabrata following in vivo exposure of the
organisms to azinphos-methyl has already been observed. In the present
study, it was investigated whether the inter-species difference in
sensitivity to cholinesterase inhibition was due to: (a) differences in the
uptake and elimination rates of the pesticide, or (b) differences in the in
vitro sensitivity of the cholinesterases to de oxon derivative. Employing a
first order mono compartmental model, it was found that the uptake and
elimination rates were very fast in both species, with very similar
bioconcentration factors. In contrast, a concentration of
azinphos-methyl-oxon 1500 times higher was needed to inhibit 50%
cholinesterase activity in the gastropod than in the oligochaete extracts.
These results indicate that the different sensitivity of both species to
cholinesterase inhibition would be more related to toxicodynamic than to
toxicokinetic processes.
Keywords: azinphos-methyl, invertebrates, toxicokinetics, cholinesterases.
Introducción
Los pesticidas organofosforados se utilizan en la agricultura moderna en
todo el mundo. Aunque estos pesticidas persisten en el ambiente por un
período relativamente corto de tiempo, presentan una alta toxicidad aguda
que puede representar un serio peligro para los ecosistemas (1). La
toxicidad primaria aguda de los organofosforados se debe a la inhibición de
la enzima acetilcolinesterasa (AChE), una enzima crítica en la función
normal del sistema nervioso. Por ello, la evaluación de la actividad de esta
enzima ha sido uno de los criterios de valoración más utilizado para evaluar
la exposición de la vida silvestre a estos pesticidas (2).
Los organofosforados que presentan un átomo de oxígeno unido a un átomo
de fósforo son altamente reactivos y no necesitan ningún tipo de activación
para inhibir la actividad de las colinesterasas (ChE). Por otro lado,
aquellos que poseen un átomo de azufre unido al átomo de fósforo, deben ser
metabólicamente activados a través de una desulfuración oxidativa a los
correspondientes oxones para ejercer su acción inhibitoria (3).
El metilazinfos (MAZ) (Figura 1) es un insecticida organofosforado del
tipo fosforoditioato que debe ser metabolizado al oxón derivado (MAZ-oxón)
para ejercer su acción anticolinesterásica. En nuestro país, este
insecticida es utilizado en el Norte de la Patagonia para el control de
Carpocapsa pomonella. Eventualmente, el MAZ alcanza, por escorrentía
superficial, a los cursos de agua ejerciendo su efecto tóxico en los
ecosistemas acuáticos (4).
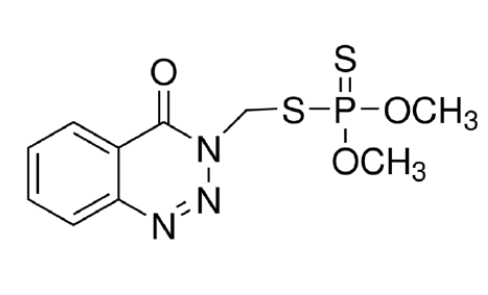
Figura 1: Estructura molecular del metilazinfos
Lumbriculus variegatus es un oligoqueto de agua dulce que ha sido
recomendado y ampliamente adoptado como un organismo estándar para estudios
de bioacumulación y ensayos de toxicidad (5,6). Biomphalaria glabrata es un
gasterópodo pulmonado que vive en aguas dulceacuícolas tropicales,
principalmente en canales, valles de irrigación y en pequeños cursos de
agua. B. glabrata es uno de los gasterópodos más estudiados por ser huésped
intermediario del parásito Schistosoma mansoni, que produce en el ser humano
la enfermedad denominada esquistosomiasis (7).
Se ha demostrado que L. variegatus es más sensible que B. glabrata a la
inhibición de la actividad ChE por exposición in vivo a MAZ. Así, la
exposición por 48 horas a 0,006 μg ml-1 de MAZ inhibió un 50% la actividad
ChE de L. variegatus mientras que para obtener el mismo grado de inhibición
en B. glabrata se necesitaron 5 μg ml-1 (8). Con el objetivo de ahondar en
los mecanismos involucrados en la diferencia de sensibilidad de ambas
especies, en el presente trabajo se procedió a: (1) calcular distintos
parámetros toxicocinéticos del MAZ (las constantes de absorción y de
eliminación, el factor de bioconcentración y el tiempo de vida media) en los
organismos expuestos in vivo por 48 h al pesticida y (2) estudiar el efecto
in vitro del metabolito biológicamente activo del MAZ, el MAZ-oxón sobre las
ChEs de L. variegatus y B. glabrata. La hipótesis de trabajo fue que la
diferencia de sensibilidad al MAZ entre L. variegatus y B. glabrata podría
deberse a diferencias en la sensibilidad del blanco molecular y/o a
diferencias en factores toxicocinéticos.
Materiales y Métodos
Reactivos
Los organofosforados MAZ Pestanal®, pureza >98%, y MAZ-oxón Supelco® y los
reactivos ioduro de acetiltiocolina (AsCh) y 5,5’-ditio-2-bis-nitrobenzoato
(DTNB) se compraron a Sigma Aldrich de Argentina S.A. El agua y el
acetonitrilo, calidad HPLC, se adquirieron a Sintorgan S.A. (Buenos Aires,
Argentina). Los otros reactivos utilizados en el trabajo fueron de grado
analítico, marca Carlo Erba, Mallinckrodt o Merck.
Organismos seleccionados
El oligoqueto L. variegatus, fue inicialmente obtenido de los cultivos del
Prof. K. Simkiss, Universidad de Reading, Reino Unido. Una vez en el
laboratorio, los organismos se cultivaron en peceras de plástico de 12
litros con aireación como se describe en (9). Para todos los bioensayos se
utilizaron organismos adultos de 2,5 ± 0,5 cm de largo.
Los cultivos de B. glabrata fueron obtenidos inicialmente del Laboratorio de
Invertebrados, Departamento de Ciencias Biológicas, Facultad de Ciencias
Exactas y Naturales, UBA. Los organismos fueron luego cultivados en nuestros
laboratorios en peceras de vidrio aireadas (9). En todos los experimentos,
se emplearon organismos adultos de tamaño similar (18 ± 2 mm de diámetro).
Para los cultivos y los bioensayos se utilizó agua de red, declorada por al
menos 24 horas, con las siguientes propiedades físico-químicas: dureza total
= 67 ± 3 mg CaCO3 L-1; alcalinidad = 29 ± 2 mg CaCO3 L-1; pH = 7,0 ± 0,2; y
conductividad = 250 ± 17 µS cm-1. Para los bioensayos, el agua declorada fue
filtrada por una columna de carbón a fin de eliminar la materia orgánica que
pudiera estar disuelta.
Ensayos de toxicocinética
Los experimentos de absorción se llevaron a cabo bajo condiciones estáticas,
sin aireación y sin suministrar alimento, en peceras de 3 L conteniendo 600
ml de solución de pesticida en agua declorada y filtrada. La concentración
de MAZ fue de 5 µg ml-1. Esta concentración se corresponde con el nivel de
exposición que produce, en B. glabrata, un 50 % de inhibición sobre la
actividad de ChEs a las 48 h (8). Las soluciones acuosas conteniendo el
pesticida se prepararon a partir de una solución concentrada de MAZ en
acetona, siendo la concentración final de acetona en las peceras de
exposición de 0,05 %. Los caracoles y las lombrices se expusieron en forma
independiente por 0, 20, 40 minutos y 1, 2, 3, 4, 5, 6 y 24 horas. Una vez
transcurrido el tiempo de exposición, cada caracol o una muestra de 10 a 15
lombrices se retiraron de las peceras, se enjuagaron con agua destilada para
eliminar cualquier resto de MAZ procedente de la solución acuosa y se
secaron con papel absorbente. En el caso de los caracoles, se les retiró el
caparazón. Las muestras se pesaron (187 ± 62 mg/caracol y 245 ± 67
mg/muestra de lombrices), se homogeneizaron en 1,5 ml de agua destilada, se
desproteinizaron y se sometieron a extracción con acetato de etilo y
análisis químico.
Para determinar los parámetros toxicocinéticos se utilizó un modelo
unicompartimental, en el cual la concentración de un xenobiótico en un
organismo acuático es descripta como el resultado de una cinética de primer
orden tanto en la absorción como en la eliminación (10):
dCorg/dt = k1.CW – k2.Corg
En esta ecuación, k1 es la constante de velocidad de absorción (ml g-1 h-1)
y k2 es la constante de velocidad de eliminación (h-1); CW es la
concentración de exposición en el agua del xenobiótico (μg ml-1) y Corg es
la concentración del compuesto en el organismo acuático (μg g-1). Frente a
una concentración de exposición constante, esta ecuación puede ser integrada
como:
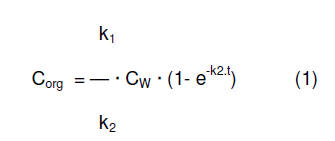
En una situación de estado estacionario (dCorg/dt = 0) esta ecuación se
puede simplificar a
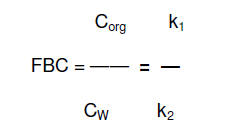
Las constantes k1 y k2
pueden determinarse a través del ajuste matemático por iteraciones múltiples
a la ecuación (1) de los datos obtenidos durante la fase de absorción.
A partir del valor de k2 se puede obtener el tiempo de vida media de un
xenobiótico, esto es, el tiempo necesario para que la concentración de dicho
compuesto descienda hasta alcanzar un valor igual a la mitad de su
concentración inicial.
t ½ = ln2/ k2 = 0,693/ k2
Separación y cuantificación del MAZ
La separación y cuantificación del MAZ en muestras de agua y en el material
biológico se realizó en un cromatógrafo líquido de alta performance,
Class-VP Shimadzu, con columna de fase reversa C18, Supelcosil LC 18, 5 μm,
con dimensiones de 25 cm por 4,6 mm, y guarda columna Supelguard LC-18 de 2
cm, bomba de flujo isocrático, detector UV-visible, perteneciente al
Laboratorio de Control de Calidad, Servicio de Farmacia, Hospital Nacional
Prof. Dr. Alejandro Posadas. Las corridas se desarrollaron en todos los
casos a una temperatura del horno de 40 °C. La columna se equilibró con una
fase móvil compuesta por acetonitrilo-agua (45:55 V/V) previamente a la
inyección de la muestra. El flujo de la corrida fue de 1ml min-1 y la
detección se realizó con un detector UV-Visible a 220 nm. La respuesta fue
lineal en el rango de concentraciones comprendido entre 0,17-54,6 μg ml-1 y
la recta de regresión lineal fue y = 118.736,73x (mg L-1) + 974,10. El
límite de cuantificación fue de 0,17 μg ml-1.
Inhibición in vitro de la actividad ChE por MAZ-oxón
Los tejidos de L. variegatus y B. glabrata fueron homogenizados en buffer
Tris/HCl 20 mM, pH 7,5, adicionado de 0,5 mM de EDTA. Los homogenatos se
centrifugaron a 11.000 x g durante 20 minutos a 4 °C. En una batería de
tubos plásticos se agregaron distintas concentraciones de MAZ-oxón disuelto
en etanol absoluto y se evaporó casi a sequedad en corriente de nitrógeno. A
estos tubos se incorporaron, al momento de efectuar la incubación, 300 μl
del sobrenadante. La incubación se realizó a 37 °C por 15 minutos.
Posteriormente se procedió a medir la actividad ChE.
La actividad ChE se midió, por duplicado, en buffer fosfato de sodio 100 mM,
pH 8,0, 0,2 mM de DTNB y 0,75 mM de AsCh (9). La actividad se registró
continuamente a 412 nm. La lectura cinética se efectuó cada 10 segundos
desde 0 a 90 segundos. Los valores se corrigieron por hidrólisis espontánea
del sustrato y por la reducción no específica del cromógeno por los
extractos titulares. La concentración de proteínas se midió por el método de
Lowry y col. (11) utilizando albúmina sérica bovina como estándar. La
actividad enzimática se expresó como μmoles de sustrato hidrolizado min-1 mg
proteína-1.
Análisis estadístico
Los resultados se expresaron como valores individuales o como el promedio ±
D.E. (desviación estándar). El efecto del MAZ-oxón sobre la actividad ChE se
analizó utilizando ANOVA de un factor y las comparaciones entre grupos se
realizaron utilizando la prueba de Tukey-Kramer. Se utilizó un nivel de
significación de 0,05.
Resultados
Toxicocinética del MAZ
Las Figuras 2 y 3 representan la cinética de absorción del metil azinfos
por L. variegatus y B. glabrata, respectivamente, durante una exposición
constante a 5 μg ml-1 del pesticida. Los datos experimentales se ajustaron
mediante un análisis de regresión no lineal iterativo a la ecuación Co=
k1/k2 Cw (1-e –k2t) y se calcularon los valores de k1 y k2. La concentración
del metil azinfos en el agua, fue constante y comprobada experimentalmente
(5,00 ± 0,10 μg ml-1) durante la exposición.
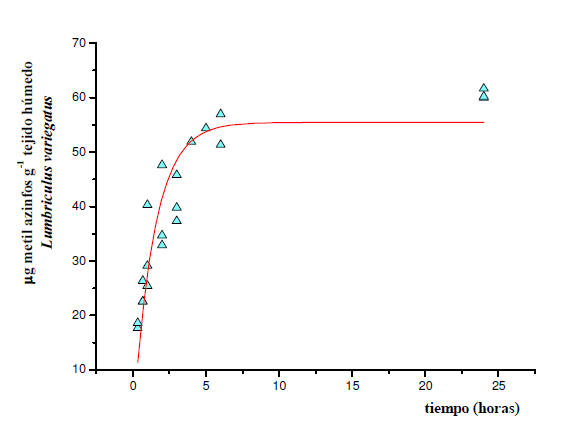
Figura 2: Cinética de absorción del MAZ por L. variegatus y el aparente
estado estacionario alcanzado. Los datos experimentales corresponden a la
fase de exposición al pesticida (5 μg ml-1). Los mismos se ajustaron
mediante un análisis de regresión no lineal iterativo a la ecuación Co=
k1/k2 Cw (1-e –k2t) utilizando el programa OriginPro versión 7.5. Cada punto
representa el contenido del compuesto parental correspondiente a un pool de
10-15 lombrices. La línea continua muestra el ajuste al modelo con un
coeficiente de determinación, R2 = 0,82179.
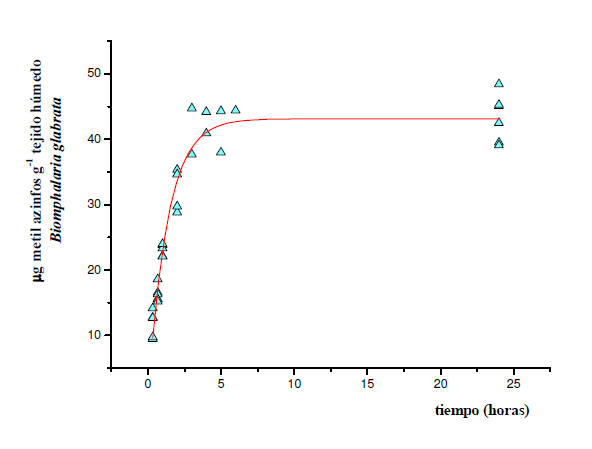
Figura 3: Cinética de absorción del MAZ por B. glabrata y el aparente estado
estacionario alcanzado. Los datos experimentales corresponden a la fase de
exposición al pesticida (5 μg ml-1). Los mismos se ajustaron mediante un
análisis de regresión no lineal iterativo a la ecuación Co= k1/k2 Cw (1-e
–k2t) utilizando el programa OriginPro versión 7.5. Cada punto representa el
contenido del compuesto parental correspondiente a un caracol. La línea
continua muestra el ajuste al modelo con un coeficiente de determinación, R2
= 0,93568.
En el caso de L. variegatus, los resultados obtenidos fueron para la
constante de absorción, k1 = 7,71 ± 0,35 ml g-1 h-1 y, para la constante de
eliminación, k2 = 0,70 ± 0,10 h-1. Según este modelo el estado estacionario
se alcanzó aproximadamente a las 5 horas de exposición. El FBCcin resultó de
11,0 ± 0,5 ml g-1 y el t ½ de 59 minutos.
En el caso de B. glabrata, los resultados obtenidos fueron para la
constante de absorción, k1 = 6,56 ± 0,13 ml g-1 h-1 y, para la constante de
eliminación, k2 = 0,76 ± 0,04 h-1. Según este modelo el estado estacionario
se alcanzó aproximadamente a las 4 horas de exposición. El FBCcin resultó de
8,6 ± 0,2 ml g-1 y el t ½ de 55 minutos.
Inhibición in vitro de la actividad ChE por MAZ-oxón
Los valores de actividad ChE en el sobrenadante de los homogenatos de L.
variegatus y B. glabrata incubados durante 15 minutos con distintas
concentraciones de MAZ-oxón, se muestran en las Figuras 4 y 5,
respectivamente. En ambos casos se observó una inhibición enzimática
dependiente de la concentración de MAZ-oxón. La concentración de xenobiótico
requerida para causar in vitro un 50% de la inhibición de la actividad ChE
(CI50) en L. variegatus y el intervalo de confianza del 95% resultó ser de
0,0025 (0,0020-0,0030) μg ml-1. En el caso de B. glabrata la CI50 y el
intervalo de confianza del 95% fueron: 3,8 (2,2-10,7) μg ml-1.
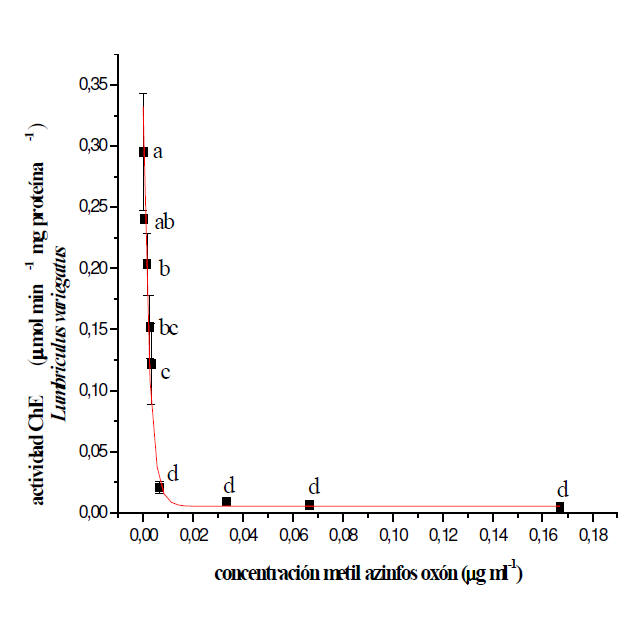
Figura 4: Efecto de concentraciones crecientes de MAZ-oxón sobre la
actividad de colinesterasa (ChE) en L. variegatus. Cada punto representa el
promedio ± D.E. de tres o más determinaciones. Las diferentes letras señalan
diferencias estadísticas significativas a p < 0,05
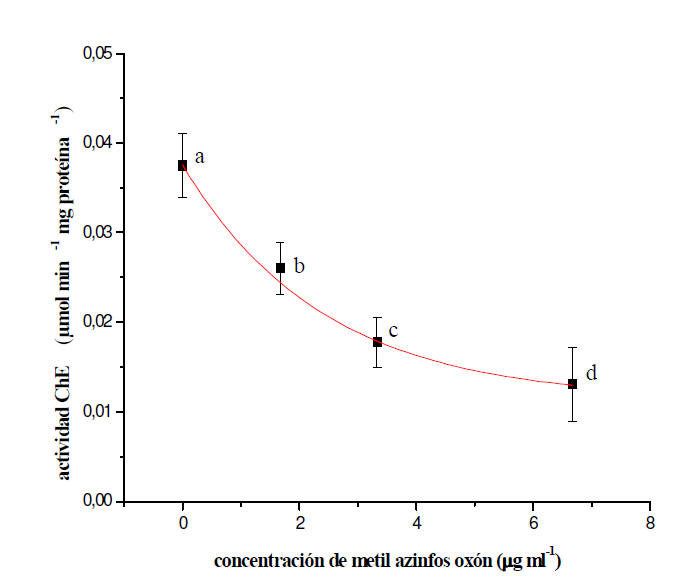
Figura 5: Efecto de concentraciones crecientes de MAZ-oxón sobre la
actividad colinesterasa (ChE) en tejidos blandos de B. glabrata. Cada punto
representa el promedio ± D.E. de tres o más determinaciones. Las diferentes
letras señalan diferencias estadísticas significativas a p < 0,05
Discusión
Estudios previos realizados exponiendo a L. variegatus y B. glabrata por
48 horas a MAZ habían permitido demostrar que los oligoquetos eran unas 1000
veces más sensibles que los gasterópodos a la inhibición in vivo de la
actividad ChE por este organofosforado (8). Los resultados del presente
trabajo indican que tanto en L. variegatus como en B. glabrata expuestos a
la concentración de 5 μg ml-1 de MAZ la absorción fue rápida, alcanzándose
un estado estacionario aparente entre las 4 y 5 horas de exposición. En
ambas especies, la velocidad de eliminación también fue rápida con un t ½
menor a 1 hora. Por último, los FBCcin calculados utilizando el modelo
unicompartimental tampoco difirieron significativamente entre ambas
especies. Estos resultados estarían indicando que la diferencia de
sensibilidad entre los oligoquetos y los gasterópodos no se debería a
diferencias en sus respectivas cinéticas de absorción y/o eliminación del
MAZ ni en su capacidad de bioconcentrarlo.
Además de procesos toxicocinéticos, procesos toxicodinámicos tales como
diferencias en la sensibilidad de los enzimas blanco pueden contribuir a la
diferente susceptibilidad para la inhibición de las ChEs entre las especies.
En el presente trabajo se pudo establecer que la actividad ChE de L.
variegatus fue 1500 veces más sensible al MAZ-oxón que la de B. glabrata.
Estos resultados podrían deberse, por un lado, a las diferencias en las ChEs
presentes en ambos organismos, considerando que trabajos previos han
demostrado que la actividad principal en L. variegatus es
butirilcolinesterasa y en B. glabrata es AChE (8). Por otro lado, también
podrían deberse a la presencia en los extractos de carboxilesterasas con
distinta afinidad por el MAZ-oxón. Estas enzimas juegan un rol importante en
el metabolismo y subsecuente detoxificación de muchos pesticidas y drogas.
Las carboxilesterasas participan en la detoxificación de los
organofosforados de dos maneras: a) por hidrólisis de uniones éster de
aquellos organofosforados que contienen este tipo de unión y b) por unión
estequiométrica de los organofosforados en el sitio activo de la enzima
produciendo su inhibición. Debido a esta unión estequiométrica, varios
investigadores han postulado que las carboxilesterasas protegerían a la AChE
de la toxicidad mediada por estos pesticidas (12, 13). Así, la presencia en
B. glabrata de carboxilesterasas mucho más sensibles al MAZ-oxón que las de
L. variegatus podrían contribuir, al menos en parte, a explicar por qué se
necesita una mayor concentración de MAZ-oxón para inhibir la actividad ChE
de B. glabrata. En este sentido, estudios in vivo mostraron que mientras las
carboxilesterasas de B. glabrata que utilizan p-nitrofenilbutirato como
sustrato son más sensibles que las ChEs a la inhibición por MAZ, en el caso
de L. variegatus ocurre lo contrario (14,15).
Referencias
1. Laguerre C., Sanchez-Hernandez, J., Kohler, H.R., Triebskorn, R.,
Capowiez, Y., Rault, M., Mazzia, C. (2009). B-type esterases in the snail
Xeropicta derbentina: An enzymological analysis to evaluate their use as
biomarkers of pesticide exposure. Environ. Poll. 157:199-207.
2. Sanchez-Hernandez, J.C. (2007). Ecotoxicological Perspectives of
B-Esterases in the Assessment of Pesticide Contamination. In: Plattenberg,
R. H. (Ed.), Environmental Pollution: New Resarch. Nova Science Publishers,
Inc. N. Y., USA, pp. 1-45.
3. Gupta, R. C.(2006). Classification and Uses of Organophosphates and
Carbamates. In Gupta, R. C.(Ed), Toxicology of Organophosphate and Carbamate
Compounds. Elesevier Academic Press: Burlington, MA, pp 5-24.
4. Loewy, R.M., Monza, L.B., Kiers, V.E., Savini, M.C. (2011). Pesticide
distribution in an agricultural environment in Argentina. J. Environ. Sci.
Health B. 46: 662-670.
5. USEPA, United States Environmental Protection Agency (2000). Methods for
measuring the toxicity and bioaccumulation of sediment-associated
contaminants with freshwater invertebrates, 600/R-94/024. US EPA Washington,
DC.
6. ASTM, American Society for Testing and Materials (1995). Standard guide
for conducting sediment toxicity tests with freshwater invertebrates. E
1393-94ª. En: Annual Book of ASTM Standards. Vol 11.05. Philadelphia, PA.
7. Balaban, A. E., Fried, B. (2013) Maintenance of Biomphalaria glabrata
infected with Schistosoma mansoni in artificial spring water, deionized
water, or conditioned tap water. Comparative Parasitology 80(1):118-119.
2013
8. Kristoff, G., Verrengia Guerrero, N., Pechén de D’Angelo, A.M., Cochón,
A.C. (2006). Inhibition of cholinesterase activity by azinphos-methyl in two
freshwater invertebrates: Biomphalaria glabrata and Lumbriculus variegatus.
Toxicology 222:185-194.
9. Kristoff, G., Verrengia Guerrero N.R., Cochón A.C. (2008).Effects of
azinphos-methyl exposure on enzymatic and non-enzymatic antioxidant defenses
in Biomphalaria glabrata and Lumbriculus variegatus. Chemosphere
72:1333-1339.
10. Nichols, J. W., Schultz, I. R., Fitzsimmons, P.N. (2006) In vitro–in
vivo extrapolation of quantitative hepatic biotransformation data for
fish. I. A review of methods, and strategies for incorporating intrinsic
clearance estimates into chemical kinetic models. Aquat. Toxicol. 78: 74–90.
11. Lowry, O.H., Rosebrough, Fan, A.L., Randall, R.J. (1951). Protein
measurement with the folin phenol reagent. J. Biol. Chem. 193: 265-275.
12. Maxwell, D. M. (1992) The specificity of carboxylesterase protection
against the toxicity of organophosphorus compounds. Toxicol. Appl.
Pharmacol. 114:306-312.
13. Wheelock, C.E., Shan, G., Ottea, J. (2005) Overview of carboxylesterases
and their role in the metabolism of insecticides. J. Pestic. Sci. 30:75-83.
14. Kristoff, G. (2010). Estudio comparativo de biomarcadores en los
invertebrados acuáticos Biomphalaria glabrata y Lumbriculus variegatus
expuestos a pesticidas de relevancia ambiental. Tesis de Doctorado. Facultad
de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires,
Argentina.
15. Kristoff, G., Chiny Barrionuevo, D., Cacciatore, L.C., Verrengia
Guerrero, N. R., Cochón, A.C. (2012). In vivo studies on inhibition and
recovery of B-esterase activities in Biomphalaria glabrata exposed to
azinphos-methyl: Analysis of enzyme, substrate and tissue dependence. Aquat.
Toxicol. 112-113, 19-26.
*Autora responsable de la publicación:
Adriana C. Cochón.Facultad de Ciencias Exactas y Naturales.
Universidad de Buenos Aires. Departamento de Química Biológica,
Ciudad Universitaria, Pab. II. 4to piso, 1428 Buenos Aires, Argentina.
TE: 0054-11-4576-3342 Fax: 0054-11-4576-3342
|
|
Revista QuímicaViva |