Curso de Introducción al Conocimiento Científico Experimental*
Capítulo 18
Enzimas
por Dra.Celia E.Coto
Introducción
Si los genes son los patrones absolutos e irreemplazables de la vida, las enzimas son los obreros especializados que hacen posible que la vida celular tenga lugar. Las enzimas son proteínas que facilitan las reacciones entre moléculas que pueden reaccionar naturalmente entre sí, pero en presencia de una enzima, la velocidad de la reacción se incrementa de tal modo que ocurre en tiempo de segundos o menos. Las enzimas son catalizadores, es decir, son sustancias que, sin consumirse en una reacción, aumentan notablemente su velocidad. No es que hagan factibles reacciones imposibles, sino que aceleran las que podrían producirse espontáneamente. Ello hace posible que, en condiciones fisiológicas, tengan lugar reacciones que sin catalizador requerirían condiciones extremas de presión, temperatura o pH. La función de las enzimas como partícipe fundamental en una reacción bioquímica fue conocida en los albores de la bioquímica clásica. En resumen, las enzimas son obreros calificados que unen, cortan, transfieren y/o modifican los grupos químicos participantes de las reacciones bioquímicas vitales para la vida.
En los capítulos 13, 14 y 15 de este curso nos referimos a las enzimas que intervienen en los procesos de síntesis de ADN y otros relacionados, pero no nos detuvimos a explicar con detalle qué son las enzimas. En este capítulo daremos un panorama general de su constitución y funciones y luego nos detendremos en el análisis de cómo si se engaña a una enzima se puede atacar una enfermedad.
¿Qué son las enzimas?
Su nombre proviene del griego énsymo (dentro de la levadura). Las enzimas son catalizadores (aumentan la rapidez) muy potentes y eficaces, químicamente son proteínas. Al igual que los catalizadores metálicos, sólo se requiere una masa pequeña para funcionar, la que se recupera indefinidamente. No llevan a cabo reacciones que sean energéticamente desfavorables, no modifican el sentido de los equilibrios químicos, sino que aceleran su consecución. Las enzimas son grandes proteínas que aceleran las reacciones químicas. En su estructura globular, se entrelazan y se pliegan mediante una o más cadenas polipeptídicas, que así aportan un pequeño grupo de aminoácidos para formar el sitio activo, o lugar donde se reconoce el sustrato, y donde se realiza la reacción. Una enzima y un sustrato no llegan a interaccionar si sus formas no encajan con exactitud. Algunos fragmentos de ARN también tienen capacidad de catalizar reacciones relacionadas con la replicación y maduración de los ácidos nucleicos, dichos fragmentos se denominan ribozimas. (1) La figura 18.1 esquematiza los pasos de un reacción enzimática en términos generales.

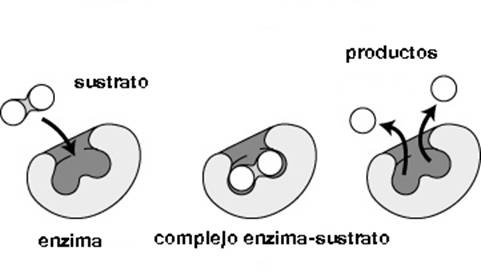
Figura 18.1: Esquematización de una reacción enzimática (tomada de la figura 7 del capítulo 13 de este curso) muestra una reacción enzimática. En el primer dibujo vemos un corte de una enzima que parecería tener forma globular que muestra su sitio activo (señalado con una flecha fina) cuya forma permite la interacción con el sustrato que debe ajustarse a la misma geometría. Es como como si calzáramos dos bolas en dos hoyos que las contengan. En este caso las dos bolas serían el sustrato que será modificado por la enzima, por ejemplo: fosforilado, hidrolizado es decir separado como en el ejemplo, etc. Como resultado final la enzima queda inalterada y sin consumir mientras que el sustrato original desapareció y se convirtió en un producto diferente.
¿Cómo funcionan?
Las enzimas son esenciales para todos los procesos biológicos ya que son las responsables de las reacciones que mantienen la vida. Cualquier mutación o disfunción en un gen responsable de la codificación de una enzima puede causar un enfermedad severa y hasta la muerte.
La acción de las enzimas se caracteriza por la formación de un complejo que representa el estado de transición. El sustrato se une a la enzima a través de numerosas interacciones débiles como ser: puentes de hidrógeno, electrostáticas, hidrófobas, etc, en un lugar específico llamado el centro activo. Este centro es una pequeña porción de la enzima, constituido por una serie de aminoácidos que interaccionan con el sustrato. Para ejercer su actividad las enzimas requieren, a menudo, de moléculas auxiliares, que se ubican en el centro activo de la enzima; en el caso de ser moléculas orgánicas reciben el nombre de coenzimas, mientras que si son iones metálicos (generalmente oligoelementos) se llaman cofactores entre los que se encuentran el hierro, cobre, yodo, manganeso, selenio, zinc, cromo, cobalto, flúor, litio y silicio. El conjunto enzima + cofactor o coenzima se denomina holoenzima, mientras que la parte proteica propiamente dicha se conoce como apoenzima. Usualmente las llamadas coenzimas no son simples moléculas auxiliares de las enzimas sino verdaderos sustratos de las reacciones pero que a diferencia del sustrato principal se regeneran fácilmente mediante reacciones simples.
Las enzimas tienen una estructura tridimensional sin la que no pueden desarrollar su actividad. En esa estructura poseen el centro activo al que se unen los sustratos y en el que se produce la reacción catalítica. Cuando el sustrato accede al centro activo, se produce un cambio en la estructura del conjunto enzima-sustrato pero una vez finalizada la catalización (catálisis: transformación química motivada por sustancias que no se alteran en el curso de la reacción) la enzima es recuperada sin ningún cambio en su estructura.
Las enzimas son esenciales para la vida ya que, de otra forma, las reacciones en las células ocurrirían lentamente.
Factores que influyen en la velocidad de las reacciones enzimáticas
Temperatura: Un aumento en la temperatura provoca un aumento de la velocidad de reacción hasta cierta temperatura óptima, ya que después de aproximadamente 450 C se comienza a producir la desnaturalización térmica. Las enzimas de muchos mamíferos tienen una temperatura óptima de 370 C, por encima de esa temperatura comienzan a inactivarse y se destruyen. Sin embargo, existen especies de bacterias y algas que habitan en fuentes de aguas termales y, en el otro extremo, ciertas bacterias árticas tienen temperaturas óptimas cercanas a 00 C. Un ejemplo interesante lo constituyen las ADN polimerasas aisladas de las bacterias que crecen en aguas termales. Cuando explicamos en el capítulo 15 la reacción de la polimerasa en cadena (PCR) comentamos que constaba de ciclos de calentamiento y enfriamento, durante el calentamiento se separaban las hebras de ADN y durante el enfriamiento se volvía a restaurar el dúplex. Lo que hizo posible esta reacción fue el descubrimiento de ADN polimerasas que funcionan a temperaturas altas.
Thermus aquaticus, denominado también Thermophilus aquaticus, es una bacteria termófila que vive en la proximidad de las fuentes de agua caliente (de 50 a 80 °C) fue descrito por Thomas Brock en 1969 en una fuente del parque de Yellowstone en Estados Unidos. Es una bacteria gram-negativa ( Gram es una tinción que permite diferenciar las bacterias según el tipo de membrana que las rodea; las gram- positivas se tiñen de violeta y las gram-negativas de rosa), aerobia (requiere oxígeno) y heterótrofa ( se alimenta de sustancias orgánicas fabricadas por otros organismos). Esta bacteria vive a temperaturas comprendidas entre 50 y 80 °C, debido a que su cóctel enzimático resiste tales condiciones. Normalmente a esas temperaturas, las proteínas constitutivas de la mayoría de los seres vivos se desnaturalizan y no vuelven a ser funcionales. Es por ello por lo que, una de sus enzimas, la ADN polimerasa de Thermus aquaticus, la llamada Taq polimerasa, es ampliamente utilizada por sus propiedades de termorresistencia en las reacciones de PCR. En efecto, esta enzima tiene una temperatura óptima de funcionamiento alrededor de los 75 °C.
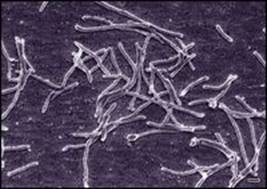
Figura 18.2. Microfotografía electrónica de Thermus aquaticus (tomada de la Wikipedia en la página dedicada a esta bacteria).
pH (grado de acidez de la solución) El pH no afecta la actividad enzimática directamente sino que modifica la concentración de protones (H+). Los protones además de alterar la estructura de la enzima y el sustrato, pueden participar también en la reacción como sustrato o producto. En esos casos, la concentración de protones afecta directamente la velocidad de la reacción. Sabiendo que las enzimas son proteínas, cualquier cambio brusco de pH, puede alterar el carácter iónico de los grupos amino (NH2-) y carboxilo (-COOH) en la superficie proteica, afectando así las propiedades catalíticas de una enzima. A pH alto o bajo se puede producir la desnaturalización de la enzima y en consecuencia su inactivación . El pH óptimo de algunas enzimas se muestra a continuación: Pepsina (1,5) ; Tripsina (7,7); Catalasa (7,6); Arginasa (9,7); Ribonucleasa (7,8)
Clasificación
El Comité de Enzimas (EC) de la Unión Internacional de Bioquímica y Biología Molecular clasifica a las enzimas en 6 clases, de acuerdo con del tipo de reacción que catalizan, se muestra en el cuadro 18.1. Se conocen al presente más de 5000 enzimas, algunas trabajan solas y otras necesitan un cofactor. Para medir la velocidad de reacción de una enzima lo que se hace es determinar la velocidad de formación del producto y la cantidad que se obtiene, la actividad se expresa como la cantidad de enzima que produce un mol de producto por minuto de reacción.
Cuadro 18.1 Clasificación de las enzimas según su función
|
Número |
Clasificación |
Propiedades bioquímicas |
|
1 |
Oxidorreductasas |
Actúan sobre muchos grupos químicos para agregar o remover átomos de hidrógeno |
|
2 |
Transferasas |
Transfieren grupos funcionales entre moléculas donantes y aceptoras. Las quinasas son transferasas especializadas que regulan el metabolismo transfiriendo fosfatos desde el ATP a otras moléculas |
|
3 |
Hidrolasas |
Agregan agua a una ligadura hidrolizándola |
|
4 |
Liasas |
Agregan agua, amoníaco o dióxido de carbono actuando sobre las dobles ligaduras o los remueven para producir enlaces dobles. |
|
5 |
Isomerasas |
Transforman ciertas sustancias en sus isómerass |
|
6 |
Ligasas |
Permiten la unión de dos moléculas con la degradación del ATP que provee la energía necesaria para que la reacción tenga lugar. |
Estructura tridimensional de las enzimas.
El avance en el conocimiento de la estructura tridimensional de las proteínas surgió mediante la tecnología que comprende primero: la cristalización de las moléculas y luego la determinación del patrón de distribución de sus átomos. Esto se logra cuando un haz de rayos X se hace atravesar el cristal, los rayos son desviados (difractados) al chocar contra los átomos de la red cristalina y se forma sobre una pantalla una figura característica. Por supuesto que sólo los expertos pueden reconstruir la forma de una molécula compleja pero lo hacen con la ayuda de programas de computación que generan estructuras en tres dimensiones. De esta forma se puede establecer cuántos polipéptidos forman la molécula, cuáles son sus plegamientos y localizar además el sitio activo si es que la molécula en cuestión es una enzima.
Consideraremos un solo ejemplo por la belleza de su estructura y su importancia vital en la duplicación y transcripción del ADN. La enzima ADN girasa o helicasa. Como ustedes recordarán de capítulos anteriores (capítulo 13) ambas hebras de un ADN se encuentran enlazadas formando una hélice. En los momentos en que se necesita copiar una de ellas, el ADN se va desenrollando por sectores, este trabajo que requiere de una máquina muy especial es la enzima ADN girasa o helicasa. La estructura de la helicasa se describió recién en 1999 y resultó ser una molécula compleja formada por seis polipéptidos individuales acomodados formando un anillo tal como se ve en la figura 18.2.
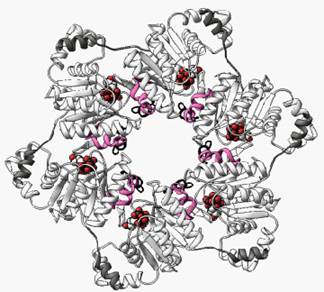
Figura 18.2 Estructura de la helicasa, se trata de una molécula compleja formada por seis helicasas individuales ensambladas de tal modo que conforman un motor en forma de anillo que desenrrolla el ADN. El motor se desplaza por el agujero central de una de las hebras y fuerza su paso a través de los pares de bases de la molécula de ADN doble. Las esferas rojas que anidan entre dos lóbulos representan las moléculas de dTTP (deoxitimidina trifosfato) que es la molécula que le provee de energía a la enzima para actuar. Los rulos rosados representan la parte de la molécula que se aferra al ADN. (2)
No todas las helicasas son hexaméricas, hay toda una familia de helicasas de estructuras más simples. A medida que la helicasa avanza a lo largo de la cadena a razón de 300 nucleótidos por segundo va quitándole, mediante hidrólisis, los fosfatos a la molécula de dTTP que es su moneda energética, ya que sabemos que tanto de este nucleótido como del ATP la célula toma energía que le permite llevar a cabo sus reacciones. En la figura 18.3 se muestra la fórmula del dTTP que al hidrolizarse y perder pirofosfato (molécula formada por dos fosfatos) cede energía.
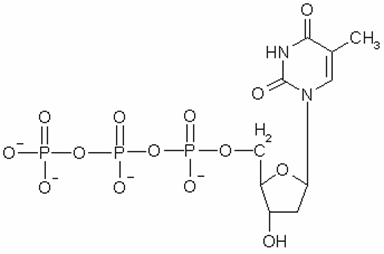
Figura 18.3. Fórmula química del dTTP.
Cómo engañar a una enzima: la base de muchos medicamentos
En una reacción típica catalizada por una enzima tanto la concentración del producto como la del reactivo son cientos de veces o miles de veces más grandes que la concentración de la enzima. La serie de eventos complejos que ocurren en esta reacción son:
E+S Û ES Þ ES* Û EP Þ E+ P
Donde ES* es un estado de transición. La cinética o velocidad de esta reacción bioquímica fue descrita por Michaelis-Menten que la formularon como ecuación y que tiene gran aplicabilidad en todas las reacciones enzimáticas bioquímicas y que permite la graficación y obtención de curvas características. La presencia en la reacción de inhibidores permite determinar en muchos casos el uso de estas sustancias como medicamentos. Para todos los interesados les recomiendo consultar la página: http://web.indstate.edu/mwking/home.html muy completa y actualizada.
Nosotros, en cambio, siguiendo la tónica de nuestro curso y completando los conocimientos impartidos en los capítulo 13,14 y 15 vamos a ver ejemplos de cómo engañar a una enzima que interviene en la síntesis de ADN virales y de ese modo conseguir moléculas antivirales eficaces. En primer lugar, vamos a analizar la acción de los análogos de nucleósidos sobre las enzimas ADN polimerasa y retrotranscriptasa.
ADN polimerasas.
La ADN polimerasa es la enzima principal involucrada en la síntesis de cadenas nuevas de ADN, esencialmente lo que hace es ir enganchando nucleótidos de adenina (A), guanina (G), citosina (C) o timina (T) unos a continuación de los otros formando una hebra larga y siguiendo el patrón de apareamiento posible que le dicta la secuencia de la cadena molde. Pero el procedimiento no es tan sencillo como lo mostramos en el capítulo 13 sino que, en realidad, para que se duplique la cadena funcionan al unísono varias enzimas debido a algunas restricciones de la propia polimerasa. Todas las ADN polimerasas conocidas (hay varias) sintetizan el ADN en la dirección 5´Þ 3´ de modo que no puede comenzar a sintetizar desde el extremo, porque sólo puede enganchar en un grupo 3´-OH y por lo tanto necesita de un cebador o primer al que pueda enganchar el primer nucleótido. Los cebadores son bases de ARN o ADN que son sintetizados por otra enzima llamada primasa. Recordemos también que, como vimos antes, se necesita una helicasa para que desenrrolle las cadenas y ofrezca a la polimerasa una cadena simple para copiar. Las ADN polimerasas son estructuras altamente conservadas lo que significa que sus subunidades catalíticas están altamente conservadas y varían poco de una especie a otra. La ADN polimerasa es una holoenzima ya que requiere ión magnesio como cofactor para funcionar adecuadamente. Algunas polimerasas tienen una función correctora esto significa que si se coloca por error un nucleótido que no corresponde al par, entonces la misma enzima lo remueve. Para ello vuelve hacia atrás un par de bases y con su actividad de exonucleasa 3’®5’ corta (separa el par erróneo) y vuelve a colocar el par correcto y continuando la replicación. Esta actividad se conoce con el de proofreading y es equivalente a corregir un texto en la prueba de galera.
Cómo se engaña a una enzima.
Las enzimas no son infalibles, pueden ser engañadas ofreciéndoles sustratos parecidos a su sustrato natural pero que posean algún cambio químico que pueda paralizar la reacción natural. Este es el principio de la obtención de muchos medicamentos contra virus que se conocen con el nombre genérico de antivirales. Así por ejemplo, si queremos combatir la infección con virus Herpes que es un ADN virus que produce esas vesículas molestas en la boca pero también que puede matar por una encefalitis, nosotros podemos evitar que se duplique su ADN dentro de la célula administrando al paciente análogos de nucleósidos. En la figura 18.4 encontramos la fórmula correspondiente a la guanosina (base normal del ADN) y en la figura 18.5 de la acicloguanosina, más conocida como aciclovir- droga efectiva contra el herpes- y del ganciclovir, droga efectiva contra el citomegalovirus (CMV).
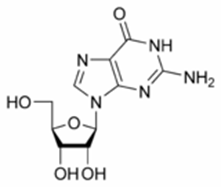
Figura 18.4 Fórmula estructural de la guanosina, resulta de la unión de la base guanina a una ribosa.
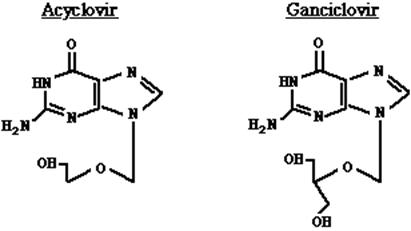
Figura 18.5. Fórmulas estructurales del aciclovir y ganciclovir. Nótese que el anillo de la ribosa está abierto y carece del OH necesario para que se enganche el próximo nucleótido.
Cuando la ADN polimerasa enganche una guanidina en la cadena que se está sintetizando porque tiene que aparearse a una citosina lo hará pero la cadena quedará cortada ya que el próximo nucleótido no tendrá dónde hacerlo, de este modo, la cadena viral no podrá ser sintetizada y no se formarán las partículas virales.
Enzima transcriptasa reversa
Se denomina así a la ADN polimerasa dependiente de ARN, que utiliza al ARN como molde en lugar del ADN. Esta enzima forma parte de los llamados retrovirus entre los que se encuentra el virus causante de SIDA, el virus HIV. Normalmente transcribe la información del ARN y la convierte en un ADN complementario formándose un híbrido ARN-ADN. Luego se digiere el ARN y se duplica la cadena de ADN dando origen a un ADN doble. Como mencionamos en otros capítulos el uso de la palabra reversa se debe a que normalmente en la células el flujo de información es ADN ® ARN ® proteína, en este caso ocurre el proceso reverso ARN ® ADN. Así como se engaña a la ADN polimerasa del virus herpes se puede engañar a la transcriptasa reversa del HIV. Como se puede observar en la figura 18.6 hay moléculas análogas a la timidina, como el conocido AZT y 3 TC que al tener bloqueado el OH de la ribosa al que debe unirse el próximo nucleótido, cortan la cadena creciente de esto modo impiden la replicación viral.
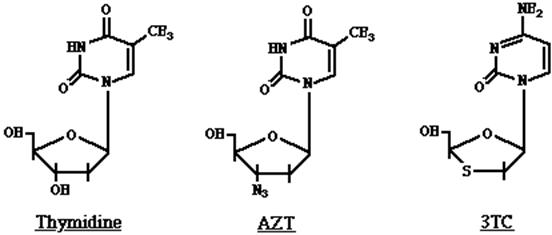
Figura 18.6. Fórmulas estructurales de timidina y análogos de timidina. En el AZT el OH está reemplazado por nitrógeno y en el 3TC por azufre.
Proteasas
Las proteasas (proteinasas, peptidasas, o enzimas proteolíticas) son enzimas que rompen las uniones peptídicas entre los aminoácidos que forman las proteínas. Recordemos que las proteínas son largos polímeros de aminoácidos con muchas funciones dentro de una célula de vital importancia como la digestión, la apoptosis, la coagulación de la sangre y otros. En la figura 18.7 se muestra la síntesis de un péptido (unión de dos aminoácidos) destacándose la unión peptídica.
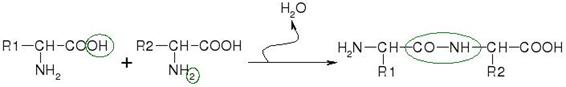
Figura 18.7 Reacción entre dos aminoácidos para dar un péptido. Como resultado de la unión se pierde una molécula de agua.
La acción de estas enzimas consiste en un proceso de ruptura proteolítica en la que usan una molécula de agua de ahí que de acuerdo a la clasificación general se las considere hidrolasas. Las proteasas pueden también servir de blancos para el ataque de microorganismos. Hay numerosas proteasas que cortan las proteínas en determinados aminoácidos, por ejemplo la familia de las caspasas de gran importancia en la apoptosis o muerte celular programada, hidrolizan la unión cuando encuentran en la cadena el aminoácido aspartato. Como esta actividad la realizan por medio de la cisteína se dice que pertenecen al grupo de las cisteín-proteasas. Otras proteasas hacen otro tipo de cortes.
Referencias
Michael W. King, Ph.D / IU School of Medicine / miking at iupui.edu
* Este curso es una contribución de Química Viva educativa (e-Lab) a la propagación del conocimiento científico entre los estudiantes de la escuela secundaria. Departamento de Química Biológica. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.
|
|
Revista QuímicaViva Revista Electrónica del Depto. de Química Biológica, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina. quimicaviva@qb.fcen.uba.ar |